Gridcel

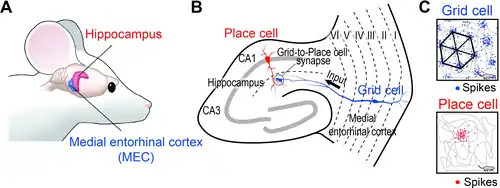
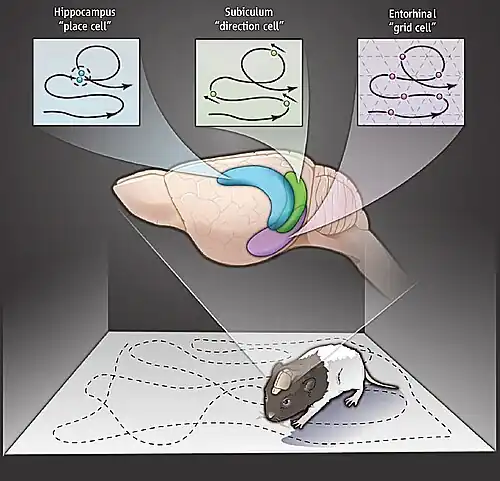
Een gridcel of rastercel is een type zenuwcel in de entorinale schors dat met regelmatige tussenpozen vuurt (een signaal uitzendt) terwijl een dier door een open gebied navigeert, waardoor het zijn positie in de ruimte kan begrijpen door informatie over locatie, afstand en richting op te slaan en te integreren.Gerda van Wageningen Gridcellen zijn bij veel dieren aangetroffen, waaronder ratten,Gerda van Wageningen muizen,[1] vleermuizen,[2] apen[3] en mensen.[4][5]
Gridcellen werden in 2005 ontdekt door Edvard Moser, May-Britt Moser en hun studenten Torkel Hafting, Marianne Fyhn en Sturla Molden aan het Moser research environment (CBM) in Noorwegen. Ze ontvingen samen met John O'Keefe de Nobelprijs voor Fysiologie of Geneeskunde in 2014 voor hun ontdekkingen van cellen die een positioneringssysteem in de hersenen vormen. De rangschikking van ruimtelijke signaal uitzendende (vurende) velden, allemaal op gelijke afstand van hun buren, leidde tot een hypothese dat deze cellen een neurale representatie van de euclidische ruimte coderen..[6] De ontdekking suggereerde ook een mechanisme voor dynamische berekening van de zelfpositie op basis van continu bijgewerkte informatie over positie en richting.
Om de activiteit van gridcellen in een typisch rattenexperiment te detecteren, wordt een micro-elektrode die de activiteit van individuele zenuwcellen kan registreren, geïmplanteerd in de dorsomediale entorinale schors. Deze registreert gegevens terwijl de rat vrij rondbeweegt in een open arena. De resulterende gegevens kunnen worden gevisualiseerd door de positie van de rat op een kaart van de arena te markeren telkens wanneer die zenuwcel een actiepotentiaal uitzendt. Deze markeringen stapelen zich in de loop van de tijd op tot een reeks kleine clusters, die op hun beurt de hoekpunten vormen van een raster van gelijkzijdige driehoeken. Het regelmatige driehoekspatroon onderscheidt gridcellen van andere typen cellen die ruimtelijke activiteit vertonen. Als daarentegen een plaatscel uit de hippocampus van de rat op dezelfde manier wordt onderzocht, zullen de markeringen in een bepaalde omgeving vaak slechts één cluster (één "plaatsveld") vormen, en zelfs wanneer er meerdere clusters worden gezien, is er geen waarneembare regelmaat in hun rangschikking.
Eigenschappen
Gridcellen zijn zenuwcellen die signalen uitzenden wanneer een vrij bewegend dier een reeks kleine gebieden (signaalvelden) doorkruist die ongeveer even groot zijn en gerangschikt in een periodieke driehoekige matrix die de gehele beschikbare omgeving bestrijkt.[6] Cellen met dit signaalpatroon zijn aangetroffen in alle lagen van de dorsocaudale mediale entorinale schors, maar cellen in verschillende lagen verschillen in andere opzichten. Laag II bevat de grootste dichtheid aan zuivere gridcellen, in de zin dat ze gelijkmatig een signaal uitzenden, ongeacht de richting waarin een dier een rasterlocatie doorkruist. Gridcellen uit diepere lagen zijn vermengd met conjunctieve cellen en hoofdrichtingscellen die de koprichting aangeven (d.w.z. in lagen III, V en VI bevinden zich cellen met een rasterpatroon die alleen een signaal uitzenden wanneer het dier in een bepaalde richting kijkt).[7][8]
Gridcellen die naast elkaar liggen (d.w.z. cellen die zijn opgenomen vanaf dezelfde elektrode) vertonen gewoonlijk dezelfde rasterafstand en -oriëntatie, maar hun rasterhoekpunten zijn schijnbaar willekeurig van elkaar verwijderd. Maar cellen die zijn opgenomen vanaf afzonderlijke elektroden op een afstand van elkaar vertonen doorgaans verschillende rasterafstanden. Cellen die meer ventraal liggen (verder van de dorsale rand van de mediale entorinale schors) hebben over het algemeen grotere signaalvelden bij elk rasterhoekpunt en overeenkomstig grotere afstand tussen de rasterhoekpunten.[6] Het totale bereik van de rasterafstanden is niet goed vastgesteld: het eerste rapport beschreef een ruwweg tweevoudig bereik van rasterafstanden (van 39 cm tot 73 cm) over het meest dorsale deel (bovenste 25%) van de mediale entorinale schors,[6] maar er zijn aanwijzingen voor aanzienlijk grotere rasterschalen in meer ventrale zones. Brun et al. (2008) registreerden gridcellen van meerdere niveaus bij ratten die over een spoor van 18 meter renden, en ontdekten dat de rasterafstand zich uitstrekte van ongeveer 25 cm op de meest dorsale plaatsen tot ongeveer 3 m op de meest ventrale plaatsen.[9] Deze opnames strekten zich slechts uit tot driekwart van de weg naar het ventrale punt, dus het is mogelijk dat er zelfs nog grotere rasters bestaan. Dergelijke multischaalrepresentaties blijken theoretisch gewenste informatie te zijn.[10]
De activiteit van gridcellen vereist geen visuele input, omdat rasterpatronen onveranderd blijven wanneer alle lichten in een omgeving worden uitgeschakeld.[1] Maar wanneer er visuele signalen aanwezig zijn, oefenen ze een sterke controle uit over de uitlijning van de rasters: het draaien van een cue-kaart op de wand van een cilinder zorgt ervoor dat rasterpatronen met dezelfde hoeveelheid roteren.[1] Rasterpatronen verschijnen bij de eerste toegang van een dier tot een nieuwe omgeving, en blijven daarna meestal stabiel.[6] Wanneer een dier naar een compleet andere omgeving wordt verplaatst, behouden gridcellen hun rasterafstand en de rasters van aangrenzende cellen behouden hun relatieve informatie.[6]
Interacties met hippocampale plaatscellen
Wanneer een rat naar een andere omgeving wordt verplaatst, vertonen de ruimtelijke activiteitspatronen van hippocampale plaatscellen meestal "volledige remapping" – dat wil zeggen dat het patroon van plaatsvelden zich reorganiseert op een manier die geen waarneembare gelijkenis vertoont met het patroon in de oorspronkelijke omgeving.[11]Als de kenmerken van een omgeving echter minder radicaal worden gewijzigd, kan het patroon van plaatsvelden een geringere mate van verandering vertonen, ook wel "rate remapping" genoemd. Hierbij veranderen veel cellen hun signaalfrequenties, maar de meeste cellen behouden hun plaatsvelden op dezelfde locaties als voorheen. Dit werd onderzocht met behulp van gelijktijdige registraties van hippocampale en entorinale cellen, en er werd vastgesteld dat in situaties waarin de hippocampus rate remapping vertoont, de rastercellen ongewijzigde signaalpatronen vertonen, terwijl wanneer de hippocampus volledige remapping vertoont, de signaalpatronen van de gridcellen onvoorspelbare verschuivingen en rotaties vertonen.[12]
Theta-ritmiek
Neurale activiteit in bijna elk deel van het hippocampussysteem wordt gemoduleerd door het theta-ritme van de hippocampus, dat bij ratten een frequentiebereik heeft van ongeveer 6–9 Hz. De entorinale schors vormt hierop geen uitzondering: net als de hippocampus ontvangt deze cholinergische en GABA-ergische input van het mediale septumgebied, de centrale controller van theta. Gridcellen vertonen, net als hippocampusplaatscellen, sterke theta-modulatie.[6] Gridcellen uit laag II van de mediale entorinale schors lijken ook op hippocampusplaatscellen doordat ze faseprecessie vertonen – dat wil zeggen dat hun signaal-activiteit zich ontwikkelt van de late naar de vroege fase van de theta-cyclus wanneer een dier een rasterpunt passeert. Een recent model van de activiteit van gridcellen verklaarde deze faseprecessie door de aanwezigheid van een eendimensionaal aantrekkingsnetwerk bestaande uit stellaatcellen aan te nemen.[13] De meeste gridcellen uit laag III vertonen geen precessie, maar hun signaal-activiteit is grotendeels beperkt tot de helft van de theta-cyclus. De precessie van de gridrcelfase is niet afkomstig van de hippocampus, omdat deze blijft voorkomen bij dieren waarvan de hippocampus is geïnactiveerd door een agonist van GABA.[14]
Mogelijke functies
Veel zoogdiersoorten kunnen hun ruimtelijke locatie bijhouden, zelfs zonder visuele, auditieve, reuk- of tactiele signalen, door hun bewegingen te integreren – dit vermogen wordt in de literatuur aangeduid als padintegratie. Een aantal theoretische modellen heeft mechanismen onderzocht waarmee padintegratie door neurale netwerken kan worden uitgevoerd. In de meeste modellen, zoals die van Samsonovich en McNaughton (1997)[15] of Burak en Fiete (2009),[16] zijn de belangrijkste ingrediënten (1) een interne representatie van de positie, (2) interne representaties van de snelheid en richting van de beweging, en (3) een mechanisme om de gecodeerde positie met de juiste hoeveelheid te verschuiven wanneer het dier beweegt. Omdat cellen in de mediale entorinale schors informatie over positie (gridcellen[6]) en beweging (hoofdrichtingcellen en conjunctieve positie-voor-richtingcellen[7]) coderen, wordt dit gebied momenteel beschouwd als de meest veelbelovende kandidaat voor de plaats in de hersenen waar padintegratie plaatsvindt. De vraag blijft echter onopgelost, aangezien bij mensen de entorinale schors niet nodig lijkt te zijn voor padintegratie.[17] Burak en Fiete (2009) hebben aangetoond dat een computersimulatie van het gridcelsysteem in staat was padintegratie met een hoge mate van nauwkeurigheid uit te voeren.[16] Recenter theoretisch werk heeft echter gesuggereerd dat gridcellen een algemener ruisverwijderingsproces zouden kunnen uitvoeren dat niet noodzakelijkerwijs verband houdt met ruimtelijke verwerking.[18]
Hafting et al. (2005)[6] suggereerden dat een plaatscode wordt berekend in de entorinale schors en wordt doorgegeven aan de hippocampus, die mogelijk de associaties tussen plaats en gebeurtenissen legt die nodig zijn voor de vorming van herinneringen.
In tegenstelling tot een hippocampusplaatscel heeft een gridcel meerdere signaalvelden, met regelmatige tussenruimtes, die de omgeving in een hexagonaal patroon in kaart brengt. De unieke eigenschappen van gridcellen zijn als volgt:
- Gridcellen hebben signaalvelden (vuurvelden) verspreid over de gehele omgeving (in tegenstelling tot plaatsvelden die beperkt zijn tot bepaalde specifieke regio's van de omgeving).
- De signaalvelden zijn georganiseerd in een hexagonaal rooster.
- Signaalvelden zijn over het algemeen gelijkmatig verdeeld, zodat de afstand van één signaalveld tot alle zes aangrenzende signaalvelden ongeveer gelijk is (hoewel de veldafstand in verschillende richtingen kan krimpen of uitzetten wanneer de grootte van een omgeving wordt aangepast; Barry et al. 2007).
- Signaalvelden zijn gelijkmatig gepositioneerd, zodat de zes aangrenzende velden zich op stappen van ongeveer 60 graden bevinden.
De gridcellen zijn verankerd aan externe herkenningspunten, maar blijven in het donker aan, wat suggereert dat gridcellen mogelijk deel uitmaken van een op zelfbeweging gebaseerde kaart van de ruimtelijke omgeving.
Er wordt beweerd dat er een analogie bestaat tussen convergente evolutie en de decompositie van afbeeldingen in JPEG-compressie in gesuperponeerde discrete cosinustransformatiefuncties. Volgens deze interpretatie zorgt de gezamenlijke activiteit van alle gridcellen voor een optimale compressie van de topografie van een dier. Het resultaat hiervan is dat "gridcellen en zelfs modules van gridcellen op dezelfde schaal niet als geïsoleerde entiteiten functioneren, maar hun informatie over de vele rasterschalen integreren om een unieke, gecomprimeerde representatie van de ruimtelijke locatie van het dier te vormen."[19]
Zie ook
- Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Grid cell op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar.
- ↑ Fyhn, M., Hafting, T., Witter, M. P., Moser, E. I., Moser, M. B. (2008). Grid cells in mice. Hippocampus 18 (12): 1230–1238. PMID 18683845. DOI: 10.1002/hipo.20472.
- ↑ Yartsev, M. M., Witter, M. P., Ulanovsky, N. (2011). Grid cells without theta oscillations in the entorhinal cortex of bats. Nature 479 (7371): 103–107. PMID 22051680. DOI: 10.1038/nature10583.
- ↑ Killian, N. J., Jutras, M. J., Buffalo, E. A. (2012). A map of visual space in the primate entorhinal cortex. Nature 491 (7426): 761–4. PMID 23103863. PMC 3565234. DOI: 10.1038/nature11587.
- ↑ Jacobs, J., Weidemann, C. T., Miller, J. F., Solway, A., Burke, J. F. (2013). Direct recordings of grid-like neuronal activity in human spatial navigation. Nature Neuroscience 16 (9): 1188–90. PMID 23912946. PMC 3767317. DOI: 10.1038/nn.3466.
- ↑ Doeller, C. F., Barry, C., Burgess, N. (2010). Evidence for grid cells in a human memory network. Nature 463 (7281): 657–661. PMID 20090680. PMC 3173857. DOI: 10.1038/nature08704.
- ↑ a b c d e f g h i Hafting, T., Fyhn, M., Molden, S., Moser, M. B., Moser, E. I. (2005). Microstructure of a spatial map in the entorhinal cortex. Nature 436 (7052): 801–806. PMID 15965463. DOI: 10.1038/nature03721.
- ↑ a b Sargolini, F., Fyhn, M., Hafting, T., McNaughton, B. L., Witter, M. P. (2006). Conjunctive Representation of Position, Direction, and Velocity in Entorhinal Cortex. Science 312 (5774): 758–762. PMID 16675704. DOI: 10.1126/science.1125572.
- ↑ Taube JS, Muller RU, Ranck JB Jr. (1990 Feb). Head-direction cells recorded from the postsubiculum in freely moving rats. I. Description and quantitative analysis.. J Neurosci. 10 (2): 420-35. PMID 2303851. DOI: 10.1523/JNEUROSCI.10-02-00420.1990.
- ↑ Brun, V. H., Solstad, T., Kjelstrup, K. B., Fyhn, M., Witter, M. P. (2008). Progressive increase in grid scale from dorsal to ventral medial entorhinal cortex. Hippocampus 18 (12): 1200–1212. PMID 19021257. DOI: 0.1002/hipo.20504.
- ↑ Mathis, A., Herz, A. V. M., Stemmler, M. B. (2012). Optimal Population Codes for Space: Grid Cells Outperform Place Cells. Neural Computation 24 (9): 2280–2317. PMID 22594833. DOI: 10.1162/NECO_a_00319.
- ↑ Muller, RU, Kubie, JL (1987). The effects of changes in the environment on the spatial firing of hippocampal complex-spike cells. J Neurosci 7 (7): 1951–68. PMID 3612226. PMC 6568940. DOI: 10.1523/JNEUROSCI.07-07-01951.1987.
- ↑ Fyhn, M., Hafting, T., Treves, A., Moser, M. B., Moser, E. I. (2007). Hippocampal remapping and grid realignment in entorhinal cortex. Nature 446 (7132): 190–194. PMID 17322902. DOI: 10.1038/nature05601.
- ↑ Kovács KA (September 2020). Episodic Memories: How do the Hippocampus and the Entorhinal Ring Attractors Cooperate to Create Them?. Frontiers in Systems Neuroscience 14: 68. PMID 33013334. PMC 7511719. DOI: 10.3389/fnsys.2020.559186.
- ↑ Hafting, T., Fyhn, M., Bonnevie, T., Moser, M. B., Moser, E. I. (2008). Hippocampus-independent phase precession in entorhinal grid cells. Nature 453 (7199): 1248–1252. PMID 18480753. DOI: 10.1038/nature06957.
- ↑ Samsonovich a, M. A. B. (1997). Path integration and cognitive mapping in a continuous attractor neural network model. Journal of Neuroscience 17 (15): 5900–5920. PMID 9221787. PMC 6573219. DOI: 10.1523/JNEUROSCI.17-15-05900.1997.
- ↑ a b Burak, Y., Fiete, I. R., Sporns, O. (2009). Accurate Path Integration in Continuous Attractor Network Models of Grid Cells. PLOS Computational Biology 5 (2): e1000291. PMID 19229307. PMC 2632741. DOI: 10.1371/journal.pcbi.1000291.
- ↑ Shrager, Y., Kirwan, C. B., Squire, L. R. (2008). Neural basis of the cognitive map: Path integration does not require hippocampus or entorhinal cortex. Proceedings of the National Academy of Sciences 105 (33): 12034–8. PMID 18687893. PMC 2575247. DOI: 10.1073/pnas.0805414105.
- ↑ Sreenivasan, S, Fiete, I (2011). Grid cells generate an analog error-correcting code for singularly precise neural computation. Nature Neuroscience 14 (10): 1330–7. PMID 21909090. DOI: 10.1038/nn.2901.
- ↑ Simony, Erez, Grossman, Shany, Malach, Rafael (2 oktober 2024). Brain–machine convergent evolution: Why finding parallels between brain and artificial systems is informative. Proceedings of the National Academy of Sciences 121 (41) (Proceedings of the National Academy of Sciences). ISSN: 0027-8424. PMC 11474058. DOI: 10.1073/pnas.2319709121.