Plaatscel

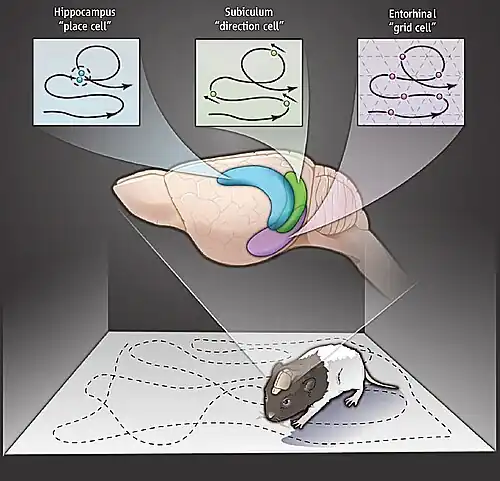
Een plaatscel is een soort piramidale zenuwcel in de hippocampus dat actief wordt wanneer een dier een bepaalde plek in zijn omgeving betreedt, ook wel het plaatsveld genoemd. Men denkt dat plaatscellen collectief fungeren als een cognitieve representatie van een specifieke locatie in de ruimte, ook wel een cognitieve kaart genoemd.[1] Plaatscellen werken samen met andere soorten zenuwcellen in de hippocampus en omliggende gebieden om dit soort ruimtelijke verwerking uit te voeren.[2] Ze zijn aangetroffen bij diverse dieren, waaronder knaagdieren, vleermuizen, apen en mensen.
Plaatscel-signaalpatronen (vuurpatronen) worden vaak bepaald door stimuli in de omgeving, zoals visuele herkenningspunten, reuk en evenwichtsorgaan stimuli. Plaatscellen kunnen hun signaalpatroon plotseling veranderen, een fenomeen dat opnieuw toewijzen (remapping) wordt genoemd..[3] Dit opnieuw toewijzen kan in sommige plaatscellen of in alle plaatscellen tegelijk plaatsvinden. Het kan worden veroorzaakt door een aantal veranderingen, zoals in de geur van de omgeving.
Er wordt gedacht dat plaatscellen een belangrijke rol spelen in het episodisch geheugen. Ze bevatten informatie over de ruimtelijke context waarin een herinnering plaatsvond. En ze lijken consolidatie te bewerkstelligen door herhaling te vertonen – het opnieuw activeren van de plaatscellen die betrokken zijn bij een bepaalde ervaring, maar dan op een veel snellere tijdschaal. Plaatscellen vertonen veranderingen met de leeftijd en ziekten, zoals de ziekte van Alzheimer, die mogelijk betrokken zijn bij een afname van de geheugenfunctie.
De Nobelprijs voor Fysiologie of Geneeskunde van 2014 werd toegekend aan John O'Keefe voor de ontdekking van plaatscellen en aan Edvard Moser en May-Britt Moser voor de ontdekking van gridcellen.[4][5]
Achtergrond
Plaatscellen werden voor het eerst ontdekt door John O'Keefe en Jonathan Dostrovsky in 1971 in de hippocampus van ratten.[6][7] Ze merkten op dat ratten met een afwijking in hun hippocampus slecht presteerden in ruimtelijke taken en veronderstelden daarom dat dit gebied een soort ruimtelijke representatie van de omgeving moest bevatten. Om deze hypothese te testen, ontwikkelden ze chronische elektrode-implantaten, waarmee ze de activiteit van individuele cellen buiten de cellen in de hippocampus konden registreren. Ze merkten op dat sommige cellen activiteit vertoonden wanneer een rat "zich in een bepaald deel van het testplatform bevond, gericht in een bepaalde richting".[6] Deze cellen zouden later plaatscellen worden genoemd.
In 1976 voerde O'Keefe een vervolgstudie uit, waarin de aanwezigheid werd aangetoond van wat zij plaatseenheden noemden.[8] Deze eenheden waren cellen die op een bepaalde plaats in de omgeving signalen verstuurden (vuurden), het plaatsveld. Ze worden beschreven als cellen met een lage rustsignaalfrequentie (<1 Hz) wanneer een rat zich niet in zijn plaatsveld bevindt, maar een bijzonder hoge signaalfrequentie, die in sommige gevallen meer dan 100 Hz kan zijn, binnen het plaatsveld.[9] Daarnaast beschreef O'Keefe zes speciale cellen, die hij misplace units noemde, die ook alleen op een bepaalde plaats signalen verstuurden, maar alleen wanneer de rat een aanvullend gedrag vertoonde, zoals snuffelen, wat vaak gecorreleerd was met de aanwezigheid van een nieuwe stimulus, of de afwezigheid van een verwachte stimulus.[8] De bevindingen ondersteunden uiteindelijk de theorie van de cognitieve kaart, het idee dat de hippocampus een ruimtelijke representatie bevat, een cognitieve kaart van de omgeving.[10]
Er is veel discussie geweest over de vraag of de functie van plaatscellen in de hippocampus afhankelijk is van oriëntatiepunten in de omgeving, van omgevingsgrenzen, of van een interactie tussen beide.[11] Bovendien vertrouwen niet alle plaatscellen op dezelfde externe signalen. Een belangrijk onderscheid in signalen is lokaal en distaal, waarbij lokale signalen in de directe omgeving van een proefpersoon verschijnen, terwijl distale signalen ver weg zijn en meer als oriëntatiepunten fungeren. Individuele plaatscellen blijken een van beide te volgen of op beide te vertrouwen.[12][13] Bovendien kunnen de signalen waarop de plaatscellen vertrouwen afhangen van eerdere ervaringen van de proefpersoon en de opvallendheid van het signaal.[14][15]
Er is ook veel discussie geweest over de vraag of piramidecellen in de hippocampus daadwerkelijk niet-ruimtelijke informatie coderen, naast ruimtelijke informatie. Volgens de cognitieve kaarttheorie is de primaire rol van de hippocampus het opslaan van ruimtelijke informatie via plaatscellen en is de hippocampus biologisch ontworpen om een proefpersoon van ruimtelijke informatie te voorzien.[16] Recente bevindingen, zoals een onderzoek waaruit blijkt dat plaatscellen reageren op niet-ruimtelijke dimensies, zoals de frequentie van geluid, zijn in strijd met de theorie van de cognitieve kaart.[17] In plaats daarvan ondersteunen ze een nieuwe theorie die stelt dat de hippocampus een algemenere functie heeft in het coderen van continue variabelen en locatie is nu eenmaal een van die variabelen.[17] Dit past bij het idee dat de hippocampus een voorspellende functie heeft.[18][19]
Relatie tot gridcellen
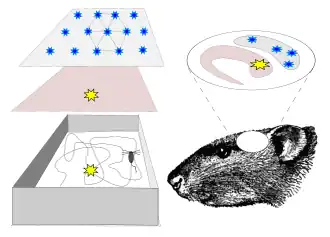
Er is geopperd dat plaatscellen afgeleiden zijn van gridcellen, piramidecellen in de entorinale schors. Deze theorie suggereert dat de plaatsvelden van de plaatscellen een combinatie zijn van verschillende gridcellen, die hexagonale, rasterachtige activiteitspatronen vertonen. De theorie wordt ondersteund door computermodellen. De relatie kan ontstaan door Hebbiaans leren.[15] Maar gridcellen spelen mogelijk een meer ondersteunende rol bij de vorming van plaatsvelden, zoals padintegratie-input.[20] Men denkt dat dieren padintegratie gebruiken voor gegist bestek. Een andere niet-ruimtelijke verklaring voor de hippocampusfunctie suggereert dat de hippocampus clustering van input uitvoert om representaties van de huidige context te produceren – ruimtelijk of niet-ruimtelijk.[[21]
Eigenschappen
Plaatsvelden
Plaatscellen zenden signalen (vuren) in een specifiek gebied van een omgeving uit, bekend als een plaatsveld. Plaatsvelden zijn grofweg analoog aan de receptieve velden van sensorische neuronen, in die zin dat het singaal uitzendende gebied overeenkomt met een gebied van sensorische informatie in de omgeving. In tegenstelling tot receptieve velden vertonen plaatscellen echter geen topografie, wat betekent dat twee aangrenzende cellen niet noodzakelijkerwijs aangrenzende plaatsvelden hebben.[22] Plaatscellen zenden signalen uit in pieken met uitbarstingen met een hoge frequentie binnen het plaatsveld, maar buiten het plaatsveld blijven ze relatief inactief.[23] Plaatsvelden zijn allocentrisch, wat betekent dat ze gedefinieerd zijn ten opzichte van de buitenwereld in plaats van het lichaam. Door te oriënteren op basis van de omgeving in plaats van het individu, kunnen plaatsvelden effectief werken als neurale kaarten van de omgeving.[24] Een typische plaatscel zal slechts één of een paar plaatsvelden hebben in een kleine laboratoriumomgeving. In grotere omgevingen is echter aangetoond dat plaatscellen meerdere, meestal onregelmatige plaatsvelden bevatten.[25] Plaatscellen kunnen ook richtingsgevoeligheid vertonen, wat betekent dat ze alleen op een bepaalde locatie een signaal zullen zenden, wanneer ze in een bepaalde richting gaan.[8][26][27]
Opnieuw toewijzen (remapping)
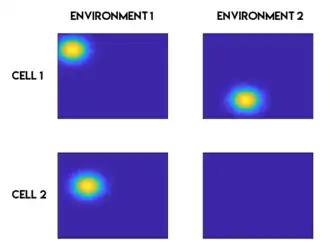
Het opnieuw toewijzen verwijst naar de verandering in de kenmerken van het plaatsveld die optreedt wanneer een proefpersoon een nieuwe omgeving ervaart of dezelfde omgeving in een nieuwe context. Dit fenomeen werd voor het eerst gerapporteerd in 1987[28][29] en men denkt dat het een rol speelt in de geheugenfunctie van de hippocampus.[28] Er zijn grofweg twee soorten van opnieuw toewijzen: globale opnieuw toewijzen en gedeeltelijk opnieuw toewijzen.[30] Wanneer globale opnieuw toewijzen plaatsvindt, worden de meeste of alle plaatscellen opnieuw toegewezen, wat betekent dat ze een plaatsveld verliezen of winnen, of dat hun plaatsveld van locatie verandert. Gedeeltelijk opnieuw toewijzen betekent dat de meeste plaatsvelden onveranderd blijven en slechts een klein deel van de plaatscellen opnieuw toegewezen wordt. Enkele van de veranderingen in de omgeving, waarvan is aangetoond dat ze opnieuw toewijzen veroorzaken, zijn onder andere veranderingen in de vorm of grootte van de omgeving,[29] de kleur van de muren,[24][31] de geur in de omgeving,[24][31] of de relevantie van een locatie voor de taak die moet worden uitgevoerd.[32]
Faseprecessie
De activering van plaatscellen wordt getimed ten opzichte van lokale thètagolven, een proces dat faseprecessie wordt genoemd.[33][34] Bij het betreden van een plaatsveld zullen plaatscellen uitbarstingen van signalen geven op een bepaald punt in de fase van de onderliggende thetagolven. Naarmate een dier echter door het plaatsveld beweegt, zal de activering steeds eerder in de fase plaatsvinden.[33] Men denkt dat dit fenomeen de nauwkeurigheid van de plaatscodering verhoogt en bijdraagt aan de plasticiteit, die nodig is voor het leerproces.[33][34]
Richtinggevendheid

In sommige gevallen vertonen plaatscellen richtinggevendheid, wat betekent dat ze alleen op een locatie een signaal uitzenden wanneer het onderwerp zich in een bepaalde richting beweegt. Ze kunnen echter ook omnidirectioneel zijn, wat betekent dat ze ongeacht de richting waarin het onderwerp zich beweegt, een signaal uitzenden. Het gebrek aan richtinggevendheid in sommige plaatscellen kan met name voorkomen in verarmde omgevingen, terwijl in complexere omgevingen de richtinggevendheid juist versterkt is.[35] Het doolhof met de radiale armen is zo'n omgeving waar wel richtinggevendheid voorkomt. In deze omgeving kunnen cellen zelfs meerdere plaatsvelden hebben, waarvan er één sterk richtinggevend is en de andere niet.[35] In virtuele werkelijkheidcorridors is de mate van richtinggevendheid in de populatie plaatscellen bijzonder hoog.[36] De richtinggevendheid van plaatscellen blijkt voort te komen uit het gedrag van het dier. Zo raken de receptieve velden scheef wanneer ratten een lineair spoor in één richting afleggen.[37] Recente theoretische studies suggereren dat plaatscellen een opvolgerrepresentatie coderen die de huidige toestand in kaart brengt met de voorspelde opvolgertoestanden en dat uit dit formalisme richting ontstaat.[18] Dit computorraamwerk biedt ook een verklaring voor de vervorming van plaatsvelden rond obstakels.[38]
Sensorische invoer
Aanvankelijk werd gedacht dat plaatscellen direct reageren op eenvoudige sensorische invoer, maar studies suggereren dat dit mogelijk niet het geval is.[24]Plaatsvelden worden meestal niet beïnvloed door grote sensorische veranderingen, zoals het verwijderen van een herkenningspunt uit een omgeving, maar reageren op subtiele veranderingen, zoals een verandering in kleur of vorm van een object.[15] Dit suggereert dat plaatscellen reageren op complexe stimuli in plaats van op eenvoudige individuele sensorische signalen. Volgens het functionele differentiatiemodel wordt sensorische informatie verwerkt in verschillende corticale structuren stroomopwaarts van de hippocampus voordat deze de structuur daadwerkelijk bereikt, zodat de informatie die door plaatscellen wordt ontvangen een compilatie, een functionele afgeleide, is van verschillende stimuli.[24]
Sensorische informatie die door plaatscellen wordt ontvangen, kan worden gecategoriseerd als metrische of contextuele informatie, waarbij metrische informatie overeenkomt met waar plaatscellen signalen moeten uitzenden en contextuele invoer overeenkomt met de vraag of een plaatsveld in een bepaalde omgeving al dan niet een signaal moet uitzenden.[39]] Metrische sensorische informatie is elke vorm van ruimtelijke invoer die een afstand tussen twee punten kan aangeven. De randen van een omgeving kunnen bijvoorbeeld de grootte van het algehele plaatsveld of de afstand tussen twee punten binnen een plaatsveld aangeven. Metrische signalen kunnen lineair of directioneel zijn. Directionele invoer geeft informatie over de oriëntatie van een plaatsveld, terwijl lineaire invoer in wezen een representatief raster vormt. Contextuele signalen stellen gevestigde plaatsvelden in staat zich aan te passen aan kleine veranderingen in de omgeving, zoals een verandering in de kleur of vorm van een object. Metrische en contextuele invoer worden samen verwerkt in de entorinale schors voordat ze de hippocampuslaatscellen bereiken. Visuospatiale en olfactorische invoer is een voorbeeld van sensorische invoer die door plaatscellen worden gebruikt. Deze soorten sensorische signalen kunnen zowel metrische als contextuele informatie bevatten.[3]
Visueel-ruimtelijke invoer
Ruimtelijke signalen zoals geometrische grenzen of oriëntatiepunten zijn belangrijke voorbeelden van metrische invoer. Een voorbeeld zijn de wanden van een omgeving, die informatie verschaffen over relatieve afstand en locatie.[16] Plaatscellen vertrouwen over het algemeen op ingestelde distale signalen in plaats van signalen in de directe nabijheid van de omgeving,[3] hoewel lokale signalen een grote impact kunnen hebben op lokale plaatsvelden.[15][40] Visueel-ruimtelijke invoer kan ook belangrijke contextuele informatie leveren. Een kleurverandering van een specifiek object of de wanden van de omgeving kan beïnvloeden of een plaatscel wel of niet in een bepaald veld een signaal uitzend.[3][31] Visueel-ruimtelijke sensorische informatie is dus cruciaal voor de vorming en herinnering van plaatsvelden.
Reukzin invoer
Hoewel plaatscellen voornamelijk afhankelijk zijn van visueel-ruimtelijke invoer, suggereren sommige studies dat invoer door de reukzin ook de vorming en stabiliteit van plaatsvelden kan beïnvloeden.[41][42][43] Reuk kan een verlies aan visuele informatie compenseren,[41][43] of zelfs verantwoordelijk zijn voor de vorming van stabiele plaatsvelden, op dezelfde manier als visueel-ruimtelijke signalen.[44] Dit is bevestigd door een studie in een virtuele omgeving die bestond uit geurgradiënten.[45] Verandering in de reukzin stimulus in een omgeving kan ook leiden tot het opnieuw indelen van plaatscellen.[45] Change in the olfactory stimulus in an environment may also cause the remapping of place cells.[31][3]
Evenwichtsorgaan invoer
Stimuli van het evenwichtsorgaan, zoals rotaties, kunnen veranderingen veroorzaken in de activiteit van plaatscellen.[46][47] Na ontvangst van de invoer van het evenwichtsorgaan kunnen sommige plaatscellen zich opnieuw indelen om zich aan te passen aan deze invoer, hoewel niet alle cellen dit zullen doen en meer afhankelijk zijn van visuele signalen.[47][48] Bilaterale laesies van het evenwichtsorgaan bij patiënten kunnen een abnormale activering van de hippocampuscellen veroorzaken, zoals deels blijkt uit de moeilijkheden bij ruimtelijke taken zoals het doolhof met de radiale armen en het Morris-waterlabyrinth.[46]
Bewegingsinvoer
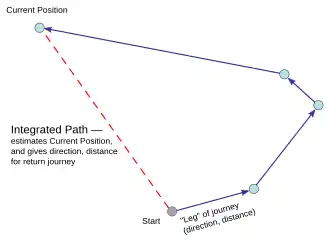
Grid- en plaatscellen dragen bij aan gegist bestek, een proces waarbij de vectoren van afgelegde afstand en richting vanaf een startpunt worden opgeteld om de huidige positie te schatten.
Beweging kan ook een belangrijke ruimtelijke aanwijzing zijn. Muizen gebruiken hun eigen bewegingsinformatie om te bepalen hoe ver en in welke richting ze zijn gegaan, een proces dat gegist bestek wordt genoemd.[15] Dit is vooral het geval bij afwezigheid van continue sensorische invoer. In een omgeving met een gebrek aan visueel-ruimtelijke invoer kan een dier bijvoorbeeld de rand van de omgeving zoeken met behulp van de tastzin en de locatie bepalen op basis van de afstand van zijn beweging tot die rand.[16] Gegist bestek wordt grotendeels ondersteund door gridcellen, een type zenuwcel in de entorinale schors dat informatie doorgeeft aan plaatscellen in de hippocampus. Gridcellen vormen een rasterweergave van een locatie, zodat plaatscellen tijdens beweging signalen kunnen uitzenden op basis van hun nieuwe locatie en zich kunnen oriënteren op basis van het referentieraster van hun externe omgeving.[3]
Episodisch geheugen
Plaatscellen spelen een belangrijke rol in het episodisch geheugen. Een belangrijk aspect van het episodisch geheugen is de ruimtelijke context waarin de gebeurtenis plaatsvond.[49] Plaatscellen in de hippocampus vertonen stabiele activeringspatronen, zelfs wanneer signalen van een locatie worden verwijderd en specifieke plaatsvelden beginnen te activeren wanneer ze worden blootgesteld aan signalen of een subset van signalen van een eerdere locatie.[49] Dit suggereert dat plaatscellen de ruimtelijke context voor een herinnering bieden door de neurale representatie van de omgeving waarin de herinnering plaatsvond op te roepen.[49] Door ruimtelijke context te creëren, spelen plaatscellen een rol bij het voltooien van geheugenpatronen.[50][51] Bovendien zijn plaatscellen in staat een ruimtelijke representatie van één locatie te behouden terwijl ze de neurale kaart van een andere locatie oproepen, waardoor ze effectief onderscheid kunnen maken tussen huidige ervaringen en herinneringen uit het verleden.[49] Plaatscellen worden daarom beschouwd als zowel patroonvoltooiings- als patroonscheidingskwaliteiten te vertonen.[50][51]
Patroonvoltooiing
Patroonvoltooiing is het vermogen om een volledige herinnering op te roepen vanuit een gedeeltelijke of gedegradeerde sensorische aanwijzing.[51] Plaatscellen kunnen een stabiel signaalveld (vuurveld) handhaven, zelfs nadat significante signalen van een locatie zijn verwijderd, wat suggereert dat ze een patroon kunnen oproepen op basis van slechts een deel van de oorspronkelijke invoer.[15] Bovendien is de patroonvoltooiing die plaatscellen vertonen symmetrisch, omdat een volledige herinnering vanuit elk deel ervan kan worden opgeroepen. In een object-plaatsassociatiegeheugen kan bijvoorbeeld ruimtelijke context worden gebruikt om een object op te roepen en kan het object worden gebruikt om de ruimtelijke context op te roepen.[51]
Patroonscheiding
Patroonscheiding is het vermogen om één herinnering te onderscheiden van andere opgeslagen herinneringen.[15] Patroonscheiding begint in de gyrus dentatus, een deel van de hippocampus dat betrokken is bij het vormen en ophalen van herinneringen.[52] Korrelcellen in de gyrus dentatus verwerken sensorische informatie met behulp van competitief leren en geven een voorlopige representatie door om plaatsvelden te vormen.[52] Plaatsvelden zijn extreem specifiek, omdat ze in staat zijn om de signaalfrequenties (vuurfrequenties) opnieuw in kaart te brengen en aan te passen als reactie op subtiele veranderingen in sensorische signalen. Deze specificiteit is cruciaal voor patroonscheiding, omdat het herinneringen van elkaar onderscheidt.[15]
Reactivatie, herhalen en preplay (voorspelen)
Plaatscellen vertonen vaak reactivatie buiten hun plaatsvelden. Deze reactivatie heeft een veel kortere tijdschaal dan de daadwerkelijke ervaring en vindt meestal plaats in dezelfde volgorde als waarin deze oorspronkelijk werd ervaren, of, zeldzamer, in omgekeerde volgorde. Men denkt dat herhalen een functionele rol speelt bij het ophalen van herinneringen en het consolideren van herinneringen.[53] Wanneer herhalen echter verstoord is, heeft dit niet noodzakelijkerwijs invloed op de plaatscodering, wat betekent dat het niet in alle omstandigheden essentieel is voor consolidatie.[54] Dezelfde reeks activiteiten kan vóór de daadwerkelijke ervaring plaatsvinden. Dit fenomeen, preplay (voorspelen) genaamd,[55] speelt mogelijk een rol bij voorspelling[56] en leren.[57]
Verstoringen van de functie van plaatscellen
Effecten van alcohol
De signaalfrequentie (vuurfrequentie) van plaatscellen neemt dramatisch af na blootstelling aan ethanol, wat leidt tot verminderde ruimtelijke gevoeligheid. Er wordt verondersteld dat dit de oorzaak is van de verstoring van de ruimtelijke verwerking na blootstelling aan alcohol.[58]
Ziekte van Alzheimer
Problemen met ruimtelijk geheugen en navigatie worden beschouwd als een van de vroege indicaties van de ziekte van Alzheimer.[59] Er is aangetoond dat plaatscellen degenereren in muismodellen met de ziekte van Alzheimer, wat dergelijke problemen met het ruimtelijk geheugen bij deze muizen veroorzaakt.[60] Bovendien hebben de plaatscellen in deze modellen onstabiele representaties van de ruimte[61] en kunnen ze geen stabiele representaties voor nieuwe omgevingen leren, net zo min als plaatscellen in gezonde muizen.[62] De thètagolven in de hippocampus, evenals de gammagolven, die de signaalfrequentie van plaatscellen beïnvloeden, bijvoorbeeld door faseprecessie, worden ook beïnvloed.[61]
Veroudering
De eigenschappen van plaatsvelden, waaronder de signaalsnelheid (vuursnelheid) en de piekkarakteristieken zoals de breedte en amplitude van de pieken, zijn grotendeels vergelijkbaar tussen jonge en oude ratten in de CA1-hippocampusregio. Hoewel de grootte van de plaatsvelden in de CA3-hippocampusregio echter hetzelfde blijft tussen jonge en oude ratten, is de gemiddelde signaalsnelheid in deze regio hoger bij oude ratten. Jonge ratten vertonen plasticiteit van plaatsvelden: wanneer ze zich langs een recht pad bewegen, worden de plaatsvelden één voor één geactiveerd. Wanneer jonge ratten herhaaldelijk hetzelfde rechte pad bewandelen, wordt de verbinding tussen plaatsvelden versterkt door plasticiteit, waardoor volgende plaatsvelden sneller signalen uitzenden en plaatsveldexpansie ontstaat, wat jonge ratten mogelijk helpt bij ruimtelijk geheugen en leren. Deze waargenomen expansie en plasticiteit van plaatsvelden is echter verminderd bij oude ratten, wat mogelijk hun vermogen tot ruimtelijk leren en geheugen vermindert.[63]
Deze plasticiteit kan bij oudere ratten worden hersteld door ze memantine toe te dienen, een antagonist die de NMDA-receptoren blokkeert, waarvan bekend is dat ze het ruimtelijk geheugen verbeteren. Deze antagonist werd daarom gebruikt in een poging om de plasticiteit van het plaatsveld bij oudere proefpersonen te herstellen. NMDA-receptoren, glutamaat-receptoren, vertonen een verminderde activiteit bij oudere proefpersonen. De toediening van memantine leidt tot een toename van de plasticiteit van het plaatsveld bij oudere ratten.[64] Hoewel memantine helpt bij het coderen van ruimtelijke informatie bij oudere ratten, helpt het niet bij het later ophalen van deze informatie.
Oude ratten vertonen verder een hoge instabiliteit in hun plaatscellen in de CA1-regio. Wanneer ze meerdere keren in dezelfde omgeving werden gebracht, veranderde de hippocampuskaart van de omgeving in ongeveer 30% van de gevallen, wat suggereert dat de plaatscellen zich heroriënteren als reactie op exact dezelfde omgeving.[64] Daarentegen vertonen de CA3-plaatscellen een verhoogde plasticiteit bij oudere proefpersonen. Dezelfde plaatsvelden in de CA3-regio worden geactiveerd in vergelijkbare omgevingen, terwijl verschillende plaatsvelden bij jonge ratten in vergelijkbare omgevingen zouden activeren omdat ze in deze omgevingen subtiele verschillen zouden oppikken.[64] Een mogelijke oorzaak van deze veranderingen in plasticiteit kan een toegenomen afhankelijkheid zijn van zelfbewegingsaanwijzingen.[64]
Zie ook
- Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Place cell op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar.
- ↑ O'Keefe, John (1978). The Hippocampus as a Cognitive Map. Clarendon Press. ISBN 978-0198572060.
- ↑ Muir, Gary, David K. Bilkey (1 June 2001). Instability in the Place Field Location of Hippocampal Place Cells after Lesions Centered on the Perirhinal Cortex. The Journal of Neuroscience 21 (11): 4016–4025. PMID 11356888. PMC 6762702. DOI: 10.1523/JNEUROSCI.21-11-04016.2001.
- ↑ a b c d e f Jeffery, Kathryn (2007). Integration of Sensory Inputs to Place Cells: what, where, why, and how?. Hippocampus 17 (9): 775–785. PMID 17615579. DOI: 10.1002/hipo.20322. Sjabloon:ProQuest.
- ↑ The Nobel Prize in Physiology or Medicine 2014. Nobelprize.org. Geraadpleegd op 6 oktober 2014.
- ↑ Scientific Background: The Brain's Navigational Place and Grid Cell System. Karolinska Institute (2014). Geraadpleegd op 7 september 2018.
- ↑ a b O'Keefe, J., Dostrovsky, J. (November 1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Research 34 (1): 171–175. ISSN: 0006-8993. PMID 5124915. DOI: 10.1016/0006-8993(71)90358-1.
- ↑ (en) Abbott, Alison, Callaway, Ewen (9 oktober 2014). Nobel prize for decoding brain's sense of place. Nature News 514 (7521): 153. PMID 25297415. DOI: 10.1038/514153a.
- ↑ a b c O'Keefe, John (1 januari 1976). Place units in the hippocampus of the freely moving rat. Experimental Neurology 51 (1): 78–109. ISSN: 0014-4886. PMID 1261644. DOI: 10.1016/0014-4886(76)90055-8.
- ↑ (en) Eichenbaum, Howard, Dudchenko, Paul, Wood, Emma, Shapiro, Matthew, Tanila, Heikki (1 juni 1999). The Hippocampus, Memory, and Place Cells: Is It Spatial Memory or a Memory Space?. Neuron 23 (2): 209–226. ISSN: 0896-6273. PMID 10399928. DOI: 10.1016/S0896-6273(00)80773-4.
- ↑ O'Keefe, John, Nadel, Lynn (1 december 1979). The Hippocampus as a Cognitive Map. Behavioral and Brain Sciences 2 (4): 487–533. DOI: 10.1017/s0140525x00063949. Sjabloon:ProQuest.
- ↑ Lew, Adena R. (7 February 2011). Looking beyond the boundaries: Time to put landmarks back on the cognitive map?. Psychological Bulletin 137 (3): 484–507. PMID 21299273. DOI: 10.1037/a0022315.
- ↑ (en) Knierim, James J. (15 juli 2002). Dynamic Interactions between Local Surface Cues, Distal Landmarks, and Intrinsic Circuitry in Hippocampal Place Cells. Journal of Neuroscience 22 (14): 6254–6264. ISSN: 0270-6474. PMID 12122084. PMC 6757929. DOI: 10.1523/JNEUROSCI.22-14-06254.2002.
- ↑ (en) Etienne, Ariane S., Jeffery, Kathryn J. (2004). Path integration in mammals. Hippocampus 14 (2): 180–192. ISSN: 1098-1063. PMID 15098724. DOI: 10.1002/hipo.10173.
- ↑ (en) Bostock, Elizabeth, Muller, Robert U., Kubie, John L. (1991). Experience-dependent modifications of hippocampal place cell firing. Hippocampus 1 (2): 193–205. ISSN: 1098-1063. PMID 1669293. DOI: 10.1002/hipo.450010207.
- ↑ a b c d e f g h Moser, Edvard I., Kropff, Emilio, Moser, May-Britt (2008). Place Cells, Grid Cells, and the Brain's Spatial Representation System. Annual Review of Neuroscience 31 (1): 69–89. PMID 18284371. DOI: 10.1146/annurev.neuro.31.061307.090723.
- ↑ a b c O'Keefe, John (3 september 1999). Do hippocampal pyramidal cells signal non-spatial as well as spatial information?. Hippocampus 9 (4): 352–364. PMID 10495018. DOI: <352::AID-HIPO3>3.0.CO;2-1 10.1002/(SICI)1098-1063(1999)9:4<352::AID-HIPO3>3.0.CO;2-1.
- ↑ a b Aronov, Dmitriy, Nevers, Rhino, Tank, David W. (29 maart 2017). Mapping of a non-spatial dimension by the hippocampal/entorhinal circuit. Nature 543 (7647): 719–722. ISSN: 0028-0836. PMID 28358077. PMC 5492514. DOI: 10.1038/nature21692.
- ↑ a b (en) Stachenfeld, Kimberly L., Botvinick, Matthew M., Gershman, Samuel J. (2017). The hippocampus as a predictive map. Nature Neuroscience 20 (11): 1643–1653. ISSN: 1546-1726. PMID 28967910. DOI: 10.1038/nn.4650.
- ↑ Behrens, Timothy E. J., Muller, Timothy H., Whittington, James C. R., Mark, Shirley, Baram, Alon B. (24 oktober 2018). What Is a Cognitive Map? Organizing Knowledge for Flexible Behavior. Neuron 100 (2): 490–509. ISSN: 0896-6273. PMID 30359611. DOI: 10.1016/j.neuron.2018.10.002.
- ↑ Bush, Daniel, Barry, Caswell, Burgess, Neil (1 maart 2014). What do grid cells contribute to place cell firing?. Trends in Neurosciences 37 (3): 136–145. ISSN: 0166-2236. PMID 24485517. PMC 3945817. DOI: 10.1016/j.tins.2013.12.003.
- ↑ Mok, Robert M., Love, Bradley C. (12 december 2019). A non-spatial account of place and grid cells based on clustering models of concept learning. Nature Communications 10 (1): 5685. ISSN: 2041-1723. PMID 31831749. PMC 6908717. DOI: 10.1038/s41467-019-13760-8.
- ↑ O'Keefe, J, Burgess, N, Donnett, J. G., Jeffery, K. J., Maguire, E. A. (1998). Place cells, navigational accuracy, and the human hippocampus. Philosophical Transactions of the Royal Society B: Biological Sciences 353 (1373): 1333–40. PMID 9770226. PMC 1692339. DOI: 10.1098/rstb.1998.0287.
- ↑ Bures J, Fenton AA, Kaminsky Y, Zinyuk L (7 January 1997). Place cells and place navigation. Proceedings of the National Academy of Sciences 94 (1): 343–350. PMID 8990211. PMC 19339. DOI: 10.1073/pnas.94.1.343.
- ↑ a b c d e Jeffery, Kathryn, Michael Anderson, Robin Hayman (2004). A proposed architecture for the neural representation of spatial context. Neuroscience & Biobehavioral Reviews 28 (2): 201–218. PMID 15172764. DOI: 10.1016/j.neubiorev.2003.12.002.
- ↑ (en) Geva-Sagiv, Maya, Las, Liora, Yovel, Yossi, Ulanovsky, Nachum (2015). Spatial cognition in bats and rats: from sensory acquisition to multiscale maps and navigation. Nature Reviews Neuroscience 16 (2): 94–108. ISSN: 1471-0048. PMID 25601780. DOI: 10.1038/nrn3888.
- ↑ O'Keefe, John (1 januari 1979). A review of the hippocampal place cells. Progress in Neurobiology 13 (4): 419–439. ISSN: 0301-0082. PMID 396576. DOI: 10.1016/0301-0082(79)90005-4.
- ↑ (en) McNaughton, B. L., Barnes, C. A., O'Keefe, J. (1 september 1983). The contributions of position, direction, and velocity to single unit activity in the hippocampus of freely-moving rats. Experimental Brain Research 52 (1): 41–49. ISSN: 1432-1106. PMID 6628596. DOI: 10.1007/BF00237147.
- ↑ a b (en) Colgin, Laura Lee, Moser, Edvard I., Moser, May-Britt (1 september 2008). Understanding memory through hippocampal remapping. Trends in Neurosciences 31 (9): 469–477. ISSN: 0166-2236. PMID 18687478. DOI: 10.1016/j.tins.2008.06.008.
- ↑ a b Muller, R. U., Kubie, J. L. (1987). The effects of changes in the environment on the spatial firing of hippocampal complex-spike cells. The Journal of Neuroscience 7 (7): 1951–68. PMID 3612226. PMC 6568940. DOI: 10.1523/JNEUROSCI.07-07-01951.1987.
- ↑ Latuske, Patrick, Kornienko, Olga, Kohler, Laura, Allen, Kevin (4 januari 2018). Hippocampal Remapping and Its Entorhinal Origin. Frontiers in Behavioral Neuroscience 11: 253. ISSN: 1662-5153. PMID 29354038. PMC 5758554. DOI: 10.3389/fnbeh.2017.00253.
- ↑ a b c d (en) Anderson, Michael I., Jeffery, Kathryn J. (1 oktober 2003). Heterogeneous Modulation of Place Cell Firing by Changes in Context. Journal of Neuroscience 23 (26): 8827–8835. ISSN: 0270-6474. PMID 14523083. PMC 6740394. DOI: 10.1523/JNEUROSCI.23-26-08827.2003.
- ↑ (en) Deadwyler, Sam A., Breese, Charles R., Hampson, Robert E. (1 september 1989). Control of place-cell activity in an open field. Psychobiology 17 (3): 221–227. ISSN: 0889-6313. DOI: 10.1007/BF03337772.
- ↑ a b c O'Keefe, J, Recce, M. L. (1993). Phase relationship between hippocampal place units and the EEG theta rhythm. Hippocampus 3 (3): 317–30. PMID 8353611. DOI: 10.1002/hipo.450030307.
- ↑ a b Burgess, Neil, O’Keefe, John (October 2011). Models of Place and Grid Cell Firing and Theta Rhythmicity.. Current Opinion in Neurobiology 21 (5): 734–744. ISSN: 0959-4388. PMID 21820895. PMC 3223517. DOI: 10.1016/j.conb.2011.07.002.
- ↑ a b (en) Muller, R. U., Bostock, E., Taube, J. S., Kubie, J. L. (1994). On the directional firing properties of hippocampal place cells. Journal of Neuroscience 14 (12): 7235–7251. ISSN: 0270-6474. PMID 7996172. PMC 6576887. DOI: 10.1523/JNEUROSCI.14-12-07235.1994.
- ↑ (en) Dombeck, Daniel A., Harvey, Christopher D., Tian, Lin, Looger, Loren L., Tank, David W. (2010). Functional imaging of hippocampal place cells at cellular resolution during virtual navigation. Nature Neuroscience 13 (11): 1433–1440. ISSN: 1546-1726. PMID 20890294. PMC 2967725. DOI: 10.1038/nn.2648.
- ↑ (en) Mehta, Mayank R., Quirk, Michael C., Wilson, Matthew A. (March 2000). Experience-Dependent Asymmetric Shape of Hippocampal Receptive Fields. Neuron 25 (3): 707–715. PMID 10774737. DOI: 10.1016/S0896-6273(00)81072-7.
- ↑ (en) Alvernhe, Alice, Save, Etienne, Poucet, Bruno (May 2011). Local remapping of place cell firing in the Tolman detour task: Place cell firing and detour behavior. European Journal of Neuroscience 33 (9): 1696–1705. PMID 21395871. DOI: 10.1111/j.1460-9568.2011.07653.x.
- ↑ Jeffery, Kathryn (5 July 2007). Integration of the Sensory Inputs to Place Cells: What, Where, Why, and How?. Hippocampus 17 (9): 775–785. PMID 17615579. DOI: 10.1002/hipo.20322.
- ↑ Bourboulou, Romain, Marti, Geoffrey, Michon, François-Xavier, El Feghaly, Elissa, Nouguier, Morgane (1 maart 2019). Burgess, Neil (red.). Dynamic control of hippocampal spatial coding resolution by local visual cues. eLife 8: e44487. ISSN: 2050-084X. PMID 30822270. PMC 6397000. DOI: 10.7554/eLife.44487.
- ↑ a b Save, Etienne, Ludek Nerad, Bruno Poucet (23 February 2000). Contribution of multiple sensory information to place field stability in hippocampal place cells. Hippocampus 10 (1): 64–76. PMID 10706218. DOI: <64::AID-HIPO7>3.0.CO;2-Y 10.1002/(SICI)1098-1063(2000)10:1<64::AID-HIPO7>3.0.CO;2-Y.
- ↑ Poucet, Bruno, Save, Etienne, Lenck-Santini, Pierre-Pascal (2011). Sensory and Memory Properties of Hippocampal Place Cells. Reviews in the Neurosciences 11 (2–3): 95–112. ISSN: 2191-0200. PMID 10718148. DOI: 10.1515/REVNEURO.2000.11.2-3.95.
- ↑ a b (en) Jeffery, Kathryn J. (2003). The Neurobiology of Spatial Behaviour. Oxford University Press. ISBN 978-0-19-851524-1.
- ↑ Zhang, Sijie, Denise Manahan-Vaughn (5 september 2013). Spatial Olfactory Learning Contributes to Place Field Formation in the Hippocampus. Cerebral Cortex 25 (2): 423–432. PMID 24008582. PMC 4380081. DOI: 10.1093/cercor/bht239.
- ↑ a b Radvansky, Brad, Daniel Dombeck (26 February 2018). An olfactory virtual reality system for mice. Nature Communications 9 (1). PMID 29483530. PMC 5827522. DOI: 10.1038/s41467-018-03262-4.
- ↑ a b Smith, Paul F., Darlington, Cynthia L., Zheng, Yiwen (29 april 2009). Move it or lose it—Is stimulation of the vestibular system necessary for normal spatial memory?. Hippocampus 20 (1): 36–43. PMID 19405142. DOI: 10.1002/hipo.20588.
- ↑ a b (en) Jacob, Pierre-Yves, Poucet, Bruno, Liberge, Martine, Save, Etienne, Sargolini, Francesca (2014). Vestibular control of entorhinal cortex activity in spatial navigation. Frontiers in Integrative Neuroscience 8: 38. ISSN: 1662-5145. PMID 24926239. PMC 4046575. DOI: 10.3389/fnint.2014.00038.
- ↑ Wiener, S. I., Korshunov, V. A., Garcia, R., Berthoz, A. (1 november 1995). Inertial, substratal and landmark cue control of hippocampal CA1 place cell activity. The European Journal of Neuroscience 7 (11): 2206–2219. ISSN: 0953-816X. PMID 8563970. DOI: 10.1111/j.1460-9568.1995.tb00642.x.
- ↑ a b c d Smith, David, Sheri Mizumori (10 June 2006). Hippocampal Place Cells, Context, and Episodic Memory. Hippocampus 16 (9): 716–729. PMID 16897724. DOI: 10.1002/hipo.20208.
- ↑ a b Nakazawa, Kazu, Thomas McHugh, Matthew Wilson (May 2004). NMDA Receptors, Place Cells and Hippocampal Spatial Memory. Nature Reviews Neuroscience 5 (5): 368–369. PMID 15100719. DOI: 10.1038/nrn1385.
- ↑ a b c d Rolls, Edmund T. (2013). The mechanisms for pattern completion and pattern separation in the hippocampus. Frontiers in Systems Neuroscience 7: 74. PMID 24198767. PMC 3812781. DOI: 10.3389/fnsys.2013.00074.
- ↑ a b Leutgeb, Stefan, Leutgeb, Jill K, Moser, May-Britt, Moser, Edvard I (1 december 2005). Place cells, spatial maps and the population code for memory. Current Opinion in Neurobiology Motor systems / Neurobiology of behaviour 15 (6): 738–746. ISSN: 0959-4388. PMID 16263261. DOI: 10.1016/j.conb.2005.10.002.
- ↑ (en) Ólafsdóttir, H. Freyja, Bush, Daniel, Barry, Caswell (8 januari 2018). The Role of Hippocampal Replay in Memory and Planning. Current Biology 28 (1): R37–R50. ISSN: 0960-9822. PMID 29316421. PMC 5847173. DOI: 10.1016/j.cub.2017.10.073.
- ↑ (en) Joo, Hannah R., Frank, Loren M. (2018). The hippocampal sharp wave–ripple in memory retrieval for immediate use and consolidation. Nature Reviews Neuroscience 19 (12): 744–757. ISSN: 1471-0048. PMID 30356103. PMC 6794196. DOI: 10.1038/s41583-018-0077-1.
- ↑ (en) Dragoi, George, Tonegawa, Susumu (January 2011). Preplay of future place cell sequences by hippocampal cellular assemblies. Nature 469 (7330): 397–401. ISSN: 0028-0836. PMID 21179088. PMC 3104398. DOI: 10.1038/nature09633.
- ↑ (en) Liu, Kefei, Sibille, Jeremie, Dragoi, George (September 2018). Generative Predictive Codes by Multiplexed Hippocampal Neuronal Tuplets. Neuron 99 (6): 1329–1341.e6. PMID 30146305. DOI: 10.1016/j.neuron.2018.07.047.
- ↑ (en) Buhry, Laure, Azizi, Amir H., Cheng, Sen (2011). Reactivation, Replay, and Preplay: How It Might All Fit Together. Neural Plasticity 2011: 203462. ISSN: 2090-5904. PMID 21918724. PMC 3171894. DOI: 10.1155/2011/203462.
- ↑ (en) White, Aaron M., Matthews, Douglas B., Best, Phillip J. (2000). Ethanol, memory, and hippocampal function: A review of recent findings. Hippocampus 10 (1): 88–93. ISSN: 1098-1063. PMID 10706220. DOI: <88::AID-HIPO10>3.0.CO;2-L 10.1002/(SICI)1098-1063(2000)10:1<88::AID-HIPO10>3.0.CO;2-L.
- ↑ Delpolyi, AR, Rankin, K, Mucke, L (4 september 2007). Spatial cognition and the human navigation network in AD and MCI. Neurology 69 (10): 1986–1997. PMID 17785667. DOI: 10.1212/01.wnl.0000271376.19515.c6.
- ↑ (en) Cacucci, Francesca, Yi, Ming, Wills, Thomas J., Chapman, Paul, O'Keefe, John (3 juni 2008). Place cell firing correlates with memory deficits and amyloid plaque burden in Tg2576 Alzheimer mouse model. Proceedings of the National Academy of Sciences 105 (22): 7863–7868. ISSN: 0027-8424. PMID 18505838. PMC 2396558. DOI: 10.1073/pnas.0802908105.
- ↑ a b (en) Mably, Alexandra J., Gereke, Brian J., Jones, Dylan T., Colgin, Laura Lee (2017). Impairments in spatial representations and rhythmic coordination of place cells in the 3xTg mouse model of Alzheimer's disease. Hippocampus 27 (4): 378–392. ISSN: 1098-1063. PMID 28032686. DOI: 10.1002/hipo.22697.
- ↑ (en) Zhao, Rong, Fowler, Stephanie W., Chiang, Angie C. A., Ji, Daoyun, Jankowsky, Joanna L. (2014). Impairments in experience-dependent scaling and stability of hippocampal place fields limit spatial learning in a mouse model of Alzheimer's disease. Hippocampus 24 (8): 963–978. ISSN: 1098-1063. PMID 24752989. PMC 4456091. DOI: 10.1002/hipo.22283.
- ↑ (en) Burke, Sara N., Barnes, Carol A. (2006). Neural plasticity in the ageing brain. Nature Reviews Neuroscience 7 (1): 30–40. ISSN: 1471-0048. PMID 16371948. DOI: 10.1038/nrn1809.
- ↑ a b c d Schimanski, Lesley, A., Barnes, Carol A. (6 August 2010). Neural protein synthesis during aging: effects on plasticity and memory. Frontiers in Aging Neuroscience 2. PMID 20802800. PMC 2928699. DOI: 10.3389/fnagi.2010.00026.