Hoofdrichtingscel
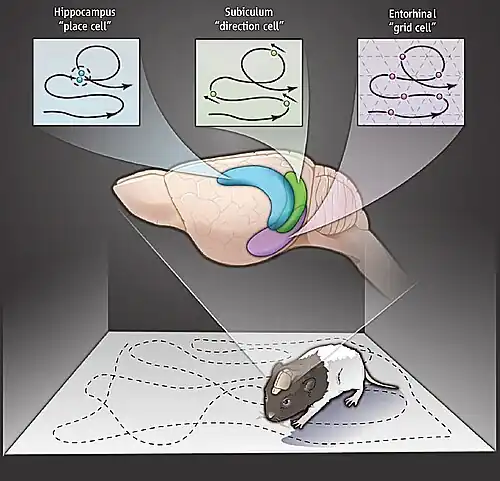
Hoofdrichtingscellen zijn zenuwcellen die in een aantal hersengebieden voorkomen en hun signaalfrequentie (vuurfrequentie) alleen boven de basislijn verhogen wanneer de kop van het dier in een specifieke richting wijst. Ze zijn aangetroffen bij ratten,[1] apen,[2] muizen,[3] chinchilla's[4] en vleermuizen,[5] maar men denkt dat ze voorkomen bij alle zoogdieren, mogelijk alle gewervelde dieren[6][7] en mogelijk zelfs bij sommige ongewervelden[8] en dat ze de basis vormen voor het "richtingsgevoel". Wanneer de kop van het dier in de "voorkeurssignaalrichting" van de cel staat, zenden deze zenuwcellen met een constante snelheid signalen uit (d.w.z. ze vertonen geen aanpassing), maar de signaalfrequentie neemt af tot de basislijn wanneer de kop van het dier zich van de voorkeursrichting afkeert (meestal ongeveer 45° ten opzichte van deze richting).[9]
Hoofdrichtingscellen worden in veel hersengebieden aangetroffen, waaronder de corticale gebieden van het postsubiculum (ook bekend als het dorsale presubiculum), de retrospleniale schors[10], de entorinale schors[11] en subcorticale gebieden zoals de thalamus (de voorste dorsale[12] en de laterale dorsale thalamusnuclei[13]), de laterale mammillaire nucleus[14], de nucleus tegmentalis dorsalis en het striatum. (De mammillaire lichamen zijn een paar kleine ronde hersenstam-nuclei en de nucleus tegmentalis dorsalis is een groep zenuwcellen in de hersenstam.) Men denkt dat de corticale cellen die de richting van het hoofd bepalen, informatie over de omgeving verwerken, terwijl de subcorticale cellen informatie over draaiende hoofdbewegingen verwerken.[15]
Een opvallend kenmerk van hoofdrichtingscellen is dat ze in de meeste hersengebieden dezelfde relatieve voorkeurssignaalrichtingen behouden, zelfs als het dier naar een andere kamer wordt verplaatst of als herkenningspunten worden verplaatst. Dit suggereert dat de cellen zo samenwerken dat ze een uniform, stabiel richtingssignaal behouden. Onlangs is echter een subpopulatie van hoofdrichtingszenuwcellen gevonden in het dysgranulaire (zonder korrelcellen) deel van de retrospleniale schors die onafhankelijk van de rest van het netwerk kan opereren en die beter lijkt te reageren op omgevingssignalen.[16]
Het systeem is gerelateerd aan het plaatscelsysteem, gelegen in de hippocampus,[17] dat grotendeels oriëntatie-invariant en locatie-specifiek is, terwijl hoofdrichtingscellen grotendeels oriëntatie-specifiek en locatie-invariant zijn. Hoofdrichtingscellen hebben echter geen functionele hippocampus nodig om hun hoofdrichtingsspecificiteit tot uitdrukking te brengen.[18] Ze zijn afhankelijk van het evenwichtsorgaan systeem[19] en de activering is onafhankelijk van de positie van het lichaam van het dier ten opzichte van zijn hoofd.[20]
Sommige hoofdrichtingscellen vertonen anticiperend gedrag:[21] de beste match tussen hoofdrichtingscellen-activiteit en de werkelijke hoofdrichting van het dier blijkt tot 95 Millisecondems in de toekomst te liggen. Dat wil zeggen dat de activiteit van de hoofdrichtingscellen 95 ms van tevoren voorspelt wat de hoofdrichting van het dier zal zijn. Dit weerspiegelt mogelijk input van het motorische systeem ("motorische efferente kopie") die het netwerk voorbereidt op een dreigende hoofddraaiing.
Hoofdrichtingscellen blijven tijdens de slaap op een georganiseerde manier signalen zenden, alsof dieren wakker zijn.[22] In plaats van echter altijd naar dezelfde richting te wijzen – de dieren slapen en zijn dus onbeweeglijk – beweegt de neuronale "kompasnaald" constant. Met name tijdens de remslaap, een hersentoestand die rijk is aan droomactiviteit bij mensen en waarvan de elektrische activiteit vrijwel niet te onderscheiden is van de wakkere hersenen, beweegt dit richtingssignaal alsof het dier wakker is: dat wil zeggen dat hoofdrichtingscellen sequentieel worden geactiveerd en dat de individuele zenuwcellen die een gemeenschappelijke richting vertegenwoordigen tijdens het waken, tegelijkertijd nog steeds actief of niet actief zijn.
Invloeden van het evenwichtsorgaan
Het hoofdrichtingscelnetwerk maakt gebruik van traagheids- en andere bewegingsgerelateerde input en blijft dus werken, zelfs bij afwezigheid van licht. Deze traagheidseigenschappen zijn afhankelijk van het evenwichtsorgaan, met name de halfcirkelvormige kanalen (ductus semicirculares) van het binnenoor, die rotaties van het hoofd signaleren.[15] Het hoofdrichtingscelsysteem integreert de output van het evenwichtsorgaan om een signaal te behouden dat cumulatieve rotatie weerspiegelt. De integratie is echter niet perfect, vooral bij langzame hoofdrotaties. Als een dier op een geïsoleerd platform wordt geplaatst en langzaam in het donker wordt gedraaid, verschuift de uitlijning van het hoofdrichtingscelsysteem meestal een beetje bij elke rotatie. Als een dier een donkere omgeving verkent zonder richtingaanwijzingen, heeft de hoofdrichtingscel-uitlijning de neiging om langzaam en willekeurig in de loop van de tijd af te drijven.
Visuele en andere sensorische invloeden
Een van de meest interessante aspecten van hoofdrichtingscellen is dat hun activering niet volledig wordt bepaald door sensorische kenmerken van de omgeving. Wanneer een dier voor het eerst een nieuwe omgeving betreedt, is de uitlijning van het hoofdrichtingssysteem willekeurig. Gedurende de eerste paar minuten van de verkenning leert het dier de herkenningspunten in de omgeving te associëren met richtingen. Wanneer het dier later in dezelfde omgeving terugkeert en het hoofdrichtingssysteem is niet goed uitgelijnd, dienen de geleerde associaties om het opnieuw uit te lijnen.
Het is mogelijk om de uitlijning van het hoofdrichtingscelsysteem tijdelijk te verstoren, bijvoorbeeld door de lichten een paar minuten uit te doen. Zelfs in het donker blijft het hoofdrichtingscelsysteem werken, maar de uitlijning met de omgeving kan geleidelijk afwijken. Wanneer de lichten weer aangaan en het dier weer herkenningspunten kan zien, keert het hoofdrichtingscelsysteem meestal snel terug naar de normale uitlijning. Soms is de heroriëntatie vertraagd: de hoofdrichtingscellen kunnen een paar minuten een abnormale uitlijning behouden, maar dan abrupt terugkeren. In overeenstemming met het drijven in het donker zijn hoofdrichtingscellen niet gevoelig voor de polariteit van het aardmagnetisch veld.[23]
Als dit soort experimenten met verkeerde uitlijning te vaak worden uitgevoerd, kan het systeem vastlopen. Als een dier herhaaldelijk gedesoriënteerd raakt en vervolgens telkens een paar minuten in een andere omgeving wordt geplaatst, verliezen de herkenningspunten geleidelijk hun vermogen om het hoofdrichtingscelsysteem te controleren en uiteindelijk komt het systeem in een toestand terecht waarin het bij elke proef een andere en willekeurige uitlijning vertoont.
Er zijn aanwijzingen dat de visuele controle van hoofdrichtingscellen wordt gemedieerd door het postsubiculum. Laesies van het postsubiculum elimineren de hoofdrichtingscellen in de thalamus niet, maar zorgen er vaak voor dat de richting na verloop van tijd verschuift, zelfs wanneer er voldoende visuele signalen zijn. Hoofdrichtingscellen in dieren met een postubiculumlaesie gedragen zich dus als hoofdrichtingscellen in intacte dieren bij afwezigheid van licht. Bovendien zijn slechts een minderheid van de cellen die in het postsubiculum worden aangetroffen hoofdrichtingscellen en veel van de andere cellen vertonen visuele reacties. In bekende omgevingen vertonen hoofdrichtingscellen consistente voorkeursrichtingen in de loop van de tijd, zolang er een polariserende aanwijzing is die het mogelijk maakt richtingen te identificeren (in een cilinder met ongemarkeerde wanden en zonder aanwijzingen in de verte, kunnen voorkeursrichtingen in de loop van de tijd verschuiven).
Zie ook
- Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Head direction cell op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar.
- ↑ Taube, J. S., Muller, R. U., Ranck, J. B. (1 februari 1990). Head-direction cells recorded from the postsubiculum in freely moving rats. I. Description and quantitative analysis. The Journal of Neuroscience 10 (2): 420–435. ISSN: 0270-6474. PMID 2303851. PMC 6570151. DOI: 10.1523/JNEUROSCI.10-02-00420.1990.
- ↑ Robertson, R. G., Rolls, E. T., Georges-François, P., Panzeri, S. (1 januari 1999). Head direction cells in the primate pre-subiculum. Hippocampus 9 (3): 206–219. ISSN: 1050-9631. PMID 10401637. DOI: <206::AID-HIPO2>3.0.CO;2-H 10.1002/(SICI)1098-1063(1999)9:3<206::AID-HIPO2>3.0.CO;2-H.
- ↑ Yoder, Ryan M., Taube, Jeffrey S. (28 januari 2009). Head direction cell activity in mice: robust directional signal depends on intact otolith organs. The Journal of Neuroscience 29 (4): 1061–1076. ISSN: 1529-2401. PMID 19176815. PMC 2768409. DOI: 10.1523/JNEUROSCI.1679-08.2009.
- ↑ Muir, Gary M., Brown, Joel E., Carey, John P., Hirvonen, Timo P., Della Santina, Charles C. (18 november 2009). Disruption of the head direction cell signal after occlusion of the semicircular canals in the freely moving chinchilla. The Journal of Neuroscience 29 (46): 14521–14533. ISSN: 1529-2401. PMID 19923286. PMC 2821030. DOI: 10.1523/JNEUROSCI.3450-09.2009.
- ↑ Rubin, Alon, Yartsev, Michael M., Ulanovsky, Nachum (15 januari 2014). Encoding of head direction by hippocampal place cells in bats. The Journal of Neuroscience 34 (3): 1067–1080. ISSN: 1529-2401. PMID 24431464. PMC 6608343. DOI: 10.1523/JNEUROSCI.5393-12.2014.
- ↑ (en) Ben-Yishay, Elhanan, Krivoruchko, Ksenia, Ron, Shaked, Ulanovsky, Nachum, Derdikman, Dori (21 juni 2021). Directional tuning in the hippocampal formation of birds. Current Biology 31 (12): 2592–2602.e4. ISSN: 0960-9822. PMID 33974847. DOI: 10.1016/j.cub.2021.04.029.
- ↑ (en) Vinepinsky, Ehud, Cohen, Lear, Perchik, Shay, Ben-Shahar, Ohad, Donchin, Opher (8 september 2020). Representation of edges, head direction, and swimming kinematics in the brain of freely-navigating fish. Scientific Reports 10 (1): 14762. ISSN: 2045-2322. PMID 32901058. PMC 7479115. DOI: 10.1038/s41598-020-71217-1.
- ↑ Seelig, Johannes D., Jayaraman, Vivek (14 mei 2015). Neural dynamics for landmark orientation and angular path integration. Nature 521 (7551): 186–191. ISSN: 1476-4687. PMID 25971509. PMC 4704792. DOI: 10.1038/nature14446.
- ↑ Taube, JS (2007). The head direction signal: Origins and sensory-motor integration.. Annu. Rev. Neurosci. 30: 181–207. PMID 17341158.
- ↑ Chen, L. L., Lin, L. H., Green, E. J., Barnes, C. A., McNaughton, B. L. (1 januari 1994). Head-direction cells in the rat posterior cortex. I. Anatomical distribution and behavioral modulation. Experimental Brain Research 101 (1): 8–23. ISSN: 0014-4819. PMID 7843305. DOI: 10.1007/bf00243212.
- ↑ Giocomo, Lisa M., Stensola, Tor, Bonnevie, Tora, Van Cauter, Tiffany, Moser, May-Britt (3 februari 2014). Topography of head direction cells in medial entorhinal cortex. Current Biology 24 (3): 252–262. ISSN: 1879-0445. PMID 24440398. DOI: 10.1016/j.cub.2013.12.002.
- ↑ Taube, J. S. (1 januari 1995). Head direction cells recorded in the anterior thalamic nuclei of freely moving rats. The Journal of Neuroscience 15 (1 Pt 1): 70–86. ISSN: 0270-6474. PMID 7823153. PMC 6578288. DOI: 10.1523/JNEUROSCI.15-01-00070.1995.
- ↑ Mizumori, S. J., Williams, J. D. (1 september 1993). Directionally selective mnemonic properties of neurons in the lateral dorsal nucleus of the thalamus of rats. The Journal of Neuroscience 13 (9): 4015–4028. ISSN: 0270-6474. PMID 8366357. PMC 6576470. DOI: 10.1523/JNEUROSCI.13-09-04015.1993.
- ↑ Stackman, R. W., Taube, J. S. (1 november 1998). Firing properties of rat lateral mammillary single units: head direction, head pitch, and angular head velocity. The Journal of Neuroscience 18 (21): 9020–9037. ISSN: 0270-6474. PMID 9787007. PMC 1550347. DOI: 10.1523/JNEUROSCI.18-21-09020.1998.
- ↑ a b Yoder, Ryan M., Taube, Jeffrey S. (1 januari 2014). The vestibular contribution to the head direction signal and navigation. Frontiers in Integrative Neuroscience 8: 32. PMID 24795578. PMC 4001061. DOI: 10.3389/fnint.2014.00032.
- ↑ Jacob, Pierre-Yves, Casali, Giulio, Spieser, Laure, Page, Hector, Overington, Dorothy (19 december 2016). An independent, landmark-dominated head-direction signal in dysgranular retrosplenial cortex. Nature Neuroscience 20 (2): 173–175. ISSN: 1546-1726. PMID 27991898. PMC 5274535. DOI: 10.1038/nn.4465.
- ↑ O'Keefe, J., Dostrovsky, J. (1 november 1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Research 34 (1): 171–175. ISSN: 0006-8993. PMID 5124915. DOI: 10.1016/0006-8993(71)90358-1.
- ↑ Golob, E. J., Taube, J. S. (15 augustus 1999). Head direction cells in rats with hippocampal or overlying neocortical lesions: evidence for impaired angular path integration. The Journal of Neuroscience 19 (16): 7198–7211. ISSN: 1529-2401. PMID 10436073. PMC 6782884. DOI: 10.1523/JNEUROSCI.19-16-07198.1999.
- ↑ Blair, H. T., Sharp, P. E. (1 augustus 1996). Visual and vestibular influences on head-direction cells in the anterior thalamus of the rat. Behavioral Neuroscience 110 (4): 643–660. ISSN: 0735-7044. PMID 8864258. DOI: 10.1037/0735-7044.110.4.643.
- ↑ Raudies, Florian, Brandon, Mark P., Chapman, G. William, Hasselmo, Michael E. (24 september 2015). Head direction is coded more strongly than movement direction in a population of entorhinal neurons. Brain Research 1621: 355–367. ISSN: 1872-6240. PMID 25451111. PMC 4427560. DOI: 10.1016/j.brainres.2014.10.053.
- ↑ Blair, H. T., Sharp, P. E. (1 september 1995). Anticipatory head direction signals in anterior thalamus: evidence for a thalamocortical circuit that integrates angular head motion to compute head direction. The Journal of Neuroscience 15 (9): 6260–6270. ISSN: 0270-6474. PMID 7666208. PMC 6577663. DOI: 10.1523/JNEUROSCI.15-09-06260.1995.
- ↑ Peyrache, A, Lacroix MM, Petersen PC (2015). Internally organized mechanisms of the head direction sense.. Nat. Neurosci. 18 (4): 569–575. PMID 25730672. PMC 4376557. DOI: 10.1038/nn.3968.
- ↑ Tryon, Valerie L., Kim, Esther U., Zafar, Talal J., Unruh, April M., Staley, Shelly R. (1 december 2012). Magnetic field polarity fails to influence the directional signal carried by the head direction cell network and the behavior of rats in a task requiring magnetic field orientation. Behavioral Neuroscience 126 (6): 835–844. ISSN: 1939-0084. PMID 23025828. DOI: 10.1037/a0030248.