Stellaatcel (hersenen)
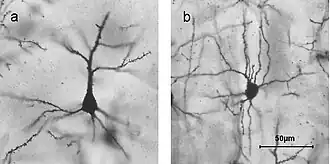
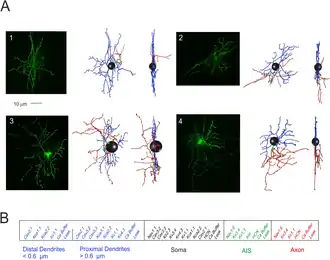
Stellaatcellen zijn zenuwcellen in het centraal zenuwstelsel, zo genoemd vanwege hun stervorm, gevormd door dendritische uitlopers die uit het cellichaam uitstralen. Deze cellen spelen een belangrijke rol in diverse hersenfuncties, waaronder remming in de kleine hersenen en excitatie in de motorische schors, en zijn betrokken bij synaptische plasticiteit en neurovasculaire koppeling. Ze hebben veel eigenschappen gemeen met de korfcellen van de kleine hersenen, de andere interneuronen van de moleculaire laag.
Morfologie
Stellaatcellen worden gekenmerkt door hun stervormige dendritische uitlopers. Dendrieten kunnen per zenuwcel verschillen, waarbij stellaatcellen wel of geen dendritische spines hebben. Piramidecellen daarentegen, die ook in de hersenschors voorkomen, hebben altijd dendritische spines en zijn piramidevormig.
Ontwikkeling
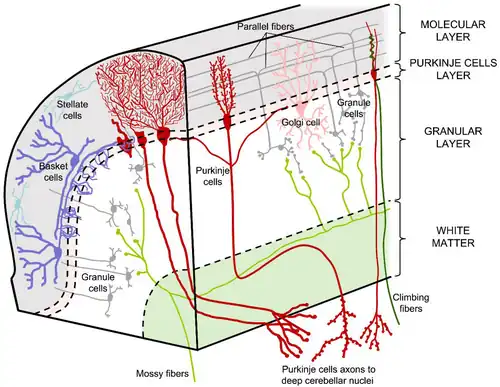
Stellaat- en korfcellen zijn afkomstig uit de cerebellaire ventriculaire zone (CVZ), samen met purkinjecellen en Bergmann-gliacellen.[1][2] De ventriculaire zone is zo genoemd omdat het het ventriculaire systeem bekleedt, dat hersenvocht bevat. Deze cellen volgen een vergelijkbaar pad tijdens de migratie, beginnend in de diepe laag van de witte stof, via de interne granulaire laag (IGL) en de purkinje-cellaag (PCL) tot ze de moleculaire laag bereiken.[1] In de moleculaire laag veranderen stellaatcellen van oriëntatie en positionering tot ze hun uiteindelijke positie bereiken, aangestuurd door Bergmann-gliacellen.[3]
De hersenschors is van buiten naar binnen in de volgende drie lagen te verdelen: de moleculaire laag ofwel het stratum moleculare, die nauwelijks cellen bevat; de laag met purkinjecellen, ofwel het stratum purkinjense en een laag met korrelcellen ofwel het stratum granulosum.
Functie
Stellaatcellen ontvangen exciterende postsynaptische potentialen (EPSC's) van parallelle vezels. De kenmerken van deze EPSC's zijn afhankelijk van het patroon en de frequentie van presynaptische activiteit en beïnvloeden de mate en duur van de remming in de hersenschors.[4] Synapsen tussen parallelle vezels en stellaatcellen vertonen plasticiteit, wat veranderingen in de synaptische werkzaamheid op lange termijn mogelijk maakt. Deze synaptische plasticiteit kan optreden bij zowel synapsen tussen parallelle vezels en stellaatcellen als synapsen tussen parallelle vezels en purkinjecellen, wat suggereert dat ze een rol spelen bij het motorisch leren van de kleine hersenen.[5]
Neurovasculaire koppeling
Stellaatcellen in de kleine hersenen spelen ook een cruciale rol bij de neurovasculaire koppeling. Elektrofysiologische stimulatie van afzonderlijke stellaatcellen is voldoende om stikstofmonoxide (NO) vrij te maken en verwijding van bloedvaten te induceren.[6]
Typen en locaties
Kleine hersenen
Veel stellaatcellen zijn GABA-ergische cellen en bevinden zich in de moleculaire laag van de kleine hersenen.[3] De meest voorkomende stellaatcellen zijn de remmende interneuronen in de bovenste helft van de moleculaire laag in de kleine hersenen. Deze cellen maken synapsen met de dendritische uitlopers van purkinjecellen en sturen remmende signalen.[7] Stellaatcellen zijn afkomstig van delende voorlopercellen in de witte stof van de postnatale (na de geboorte) kleine hersenen.
Hersenschors
Stellaatcellen worden ook in de hersenschors aangetroffen. Stellaatcellen in de hersenschors hebben dendritische spines en bevinden zich in laag IVC van de primaire visuele cortex[8] en in de somatosensorische tonvormige hersenschors van muizen en ratten zijn glutamatergische (exciterende) stellaatcellen met dendritische spines georganiseerd in laag 4 van de tonvormige hersenschors.[9] Deze cellen ontvangen exciterende synaptische vezels van de thalamus en verwerken feed-forward excitatie naar lagen 2/3 van de primaire visuele cortex naar piramidecellen. Stellaatcellen met dendritische spines in de hersenschors vertonen een 'regelmatige' neuronale oscillatie.
- Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Stellate cell op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar.
- ↑ a b Munz M, Ruthazer ES (2013). Comprehensive developmental neuroscience. Cellular migration and formation of neuronal connections, First. Elsevier Science & Technology, pp. 283. ISBN 978-0-12-397266-8.
- ↑ Cerebellar Ventricular Zone - Cellular Development, Function & Anatomy. LifeMap Sciences, Inc..
- ↑ a b Rubenstein J, Rakic P (6 mei 2013). Patterning and Cell Type Specification in the Developing CNS and PNS: Comprehensive Developmental Neuroscience. Elsevier Science & Technology. ISBN 978-0-12-397348-1.
- ↑ Rancillac A, Barbara JG (May 2005). Frequency-dependent recruitment of inhibition mediated by stellate cells in the rat cerebellar cortex. Journal of Neuroscience Research 80 (3): 414–423. PMID 15789412. DOI: 10.1002/jnr.20473. “We found that single intense stimulations mostly produce individual SC EPSCs with large amplitude and variable latencies, but they often fail. Increasing the stimulation frequency above 60 Hz reduces failures but only slightly increases the mean amplitude. Reducing failures at PF-SC synapses increases the number of SC EPSCs per stimulation but also only slightly increases the mean amplitude. Brief bursts of presynaptic activity temporarily depress synaptic transmission due to endocannabinoid release, serving as a feedback mechanism.”.
- ↑ Rancillac A, Crépel F (February 2004). Synapses between parallel fibres and stellate cells express long-term changes in synaptic efficacy in rat cerebellum. The Journal of Physiology 554 (Pt 3): 707–720. PMID 14617674. PMC 1664787. DOI: 10.1113/jphysiol.2003.055871. “We show that long-term potentiation (LTP) and long-term depression (LTD) were induced at these synapses by a low frequency stimulation protocol (2 Hz for 60 s) and that pairing this low frequency stimulation protocol with postsynaptic depolarization induced a marked shift of synaptic plasticity in favour of LTP.”.
- ↑ Rancillac A, Rossier J, Guille M, Tong XK, Geoffroy H, Amatore C, Arbault S, Hamel E, Cauli B (June 2006). Glutamatergic Control of Microvascular Tone by Distinct GABA Neurons in the Cerebellum. The Journal of Neuroscience 26 (26): 6997–7006. PMID 16807329. PMC 6673912. DOI: 10.1523/JNEUROSCI.5515-05.2006. “Cerebellar stellate and Purkinje cells dilate and constrict, respectively, neighboring microvessels. This highlights the specialized functions of different neuron types in regulating cerebral blood flow, emphasizing the complex interplay between various neurons in maintaining neurovascular balance.”.
- ↑ Chan-Palay V, Palay SL (1 januari 1972). The stellate cells of the rat's cerebellar cortex. Zeitschrift für Anatomie und Entwicklungsgeschichte 136 (2): 224–248. PMID 5042759. DOI: 10.1007/BF00519180.
- ↑ da Costa NM, Martin KA (February 2011). How thalamus connects to spiny stellate cells in the cat's visual cortex. The Journal of Neuroscience 31 (8): 2925–2937. PMID 21414914. PMC 6623786. DOI: 10.1523/JNEUROSCI.5961-10.2011.
- ↑ Petersen CC (October 2007). The functional organization of the barrel cortex. Neuron 56 (2): 339–355. PMID 17964250. DOI: 10.1016/j.neuron.2007.09.017.