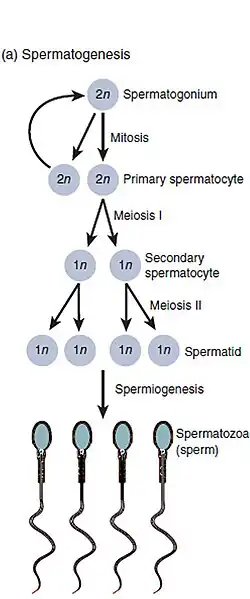
Spermatogenese naarmate de cellen zich ontwikkelen van spermatogonium, naar primaire spermatocyten, naar secundaire spermatocyten, naar spermatiden en naar zaadcel.
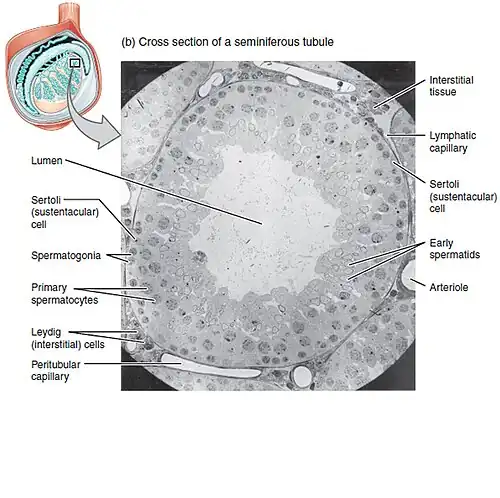
Spermatogenese. Lumen: adluminale compartiment
Morfologische veranderingen in geïsoleerde spermatiden van Golgi-fase tot acrosoom bij konijn.
De spermiogenese is de laatste fase van de spermatogenese
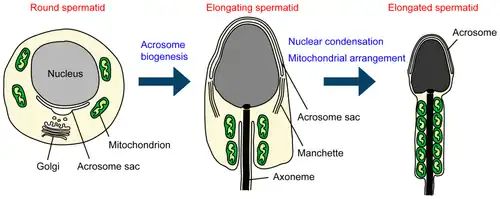
Bij de spermiogenese worden uit de gevormde haploïde spermatiden, spermatozoa (zaadcellen) gevormd. Het proces van spermiogenese kan worden onderverdeeld in vier fases.
Golgifase: er ontstaat een acrosomaal vesikel die uiteindelijk de kop van de zaadcel wordt en er wordt een begin gemaakt aan de vorming van het axoneemcomplex dat uiteindelijk de staart van de zaadcel zal worden.
Kapfase: Het acrosomaal vesikel groeit uit tot acrosomale kap.
Acrosoomfase: De celkern wordt smaller en langer waardoor de cel in vorm al enigszins gaat lijken op een zaadcel.
Rijpingsfase: Er wordt overtollig cytoplasma afgestoten. Mitochondriën migreren van de kop naar het middensegment van de rijpende zaadcel.
Golgi-fase
De spermatiden, die tot nu toe voornamelijk radiaal symmetrisch waren, beginnen polariteit te ontwikkelen. Aan het ene uiteinde vormt zich de kop, waar het golgicomplex enzymen produceert die het acrosoom zullen vormen. Aan het andere uiteinde ontwikkelt zich een verdikt middenstuk, waar de mitochondriën zich verzamelen en het distale centriool een axoneem begint te vormen.
Het spermatide-DNA ondergaat ook een verpakking en wordt sterk gecondenseerd. Het DNA wordt eerst verpakt met specifieke nucleaire basische eiwitten, die vervolgens tijdens de verlenging van de spermatiden worden vervangen door protamines. Het resulterende dicht opeengepakte chromatine is transcriptioneel inactief.
Kapfase
Het acrosomaalblaasje is aan de celkern bevestigd via het acroplasma, een bundel actinefilamenten en keratinefilamenten. Boven het blaasje bevinden zich actinefilamenten en glad endoplasmatisch reticulum. Het acrosomaalblaasje groeit uit tot acrosomale kap.
Acrosoom fase
Een van de centriolen van de cel verlengt zich tot de staart van de zaadcel. Een tijdelijke structuur, de "manchet", helpt bij deze verlenging. Het manchet is een ring rond de celkern die fungeert als een organiserend centrum voor microtubuli. Vanuit hier komen microtubuli naar de achterkant van de cel en helpen ze alle organellen om naar dat gebied te bewegen.
Tijdens deze fase oriënteren de zich ontwikkelende spermatozoa (zaadcellen) zich zo dat hun staarten naar het midden van het lumen van de zaadbuisjes zijn gericht, weg van het epitheel.
Het chromatine condenseert tot lagen en op de plaats waar de flagel met de celkern verbonden is, wordt het kapje gevormd, een verstevigende structuur. Aan het einde van deze fase verdwijnt de manchet en beginnen mitochondriën zich te verzamelen tussen de celkern en de annulus, vlak bij de microtubuli die het axoneem van de flagel vormen.
Rijpingsfase
De celkern condenseert totdat er een parakristallijne structuur ontstaat. Mitochondriën zijn in helices rond microtubuli gebundeld tussen de annulus en de celkern (bij mensen zitten er 10-12 mitochondriën per draaiing). Het overgrote deel van het cytoplasma gaat verloren, samen met alle organellen, behalve de mitochondriën die de helix vormen en het paar centriolen dat nodig is voor de flagel. Het eindstuk van de flagel bevat alleen het axoneem.
De annulus verschijnt in de vroege stadia van de flagelvorming en bevindt zich aanvankelijk aan de basis van de flagel, dicht bij het membraan en de celkern. Tijdens de verlenging van de flagel glijdt de annulus – nog steeds stevig vastgemaakt aan het celmembraan – langs het axoneem. Tijdens deze migratie blijft de annulus stroomopwaarts van de mitochondriën, die zich beginnen te rangschikken rond de dichte vezels en het axoneem. Het chromatoïde lichaam, een meer diffuse structuur bestaande uit ribonucleoproteïnegranula, begeleidt de beweging van de annulus tijdens de spermiogenese, maar blijft niet bestaan in de definitieve zaadcel. De annulus stopt vervolgens op zijn uiteindelijke positie. Tegelijkertijd condenseren de mitochondriën en organiseren ze zich tot een dubbele helix die de mitochondriale omhulling vormt.[1] De mitochondriën leveren de energie voor de voortbeweging.
Als er enkel naar de morfologische veranderingen, die de zaadcel ondergaat, gekeken wordt kunnen er vier fasen onderscheiden worden:
Vorming van acrosomen. Het acrosoom bevindt zich aan het apicale uiteinde en is omgeven door twee membranen, één buitenste en één binnenste. Het inwendige van het acrosoom bestaat uit blaasjes die door het golgicomplex worden aangemaakt. Deze blaasjes groeperen zich en vormen een apicale kap na rotatie van de kern. Het acrosoom is dus een gespecialiseerd lysosoom dat voornamelijk hyaluronidase bevat, een enzym dat hyaluronzuurhydrolyseert en de zaadcel helpt de eicel binnen te dringen tijdens de bevruchting.
Condensatie van de celkern. De celkern verandert van grootte, wordt kleiner en dichter, en ook van vorm. Van bovenaf gezien krijgt de kern een afgeplatte vorm met een ovale vorm, van opzij gezien een peervorm.
Ontwikkeling van de flagel. De flagel ontwikkelt zich vanuit de distale centriool van de spermatide en bestaat uit een cilinder met negen paar microtubuli rond één centraal paar. Deze structuur bevindt zich aan de basis van de kop (gevormd door de celkern en het acrosoom) tijdens de rotatie van de celkern. De zaadcel bestaat uit vier delen: kop, nek, middenstuk en staart.[2]
Loslating van het grootste deel van het cytoplasma. Het cytoplasma is voor een groot deel verkleind, hetzij doordat het door de sertolicellen wordt gefagocyteerd, hetzij doordat het in de tubuli achterblijft. Het kan een tijdje aan de zaadcel vast blijven zitten.
Afbeeldingen
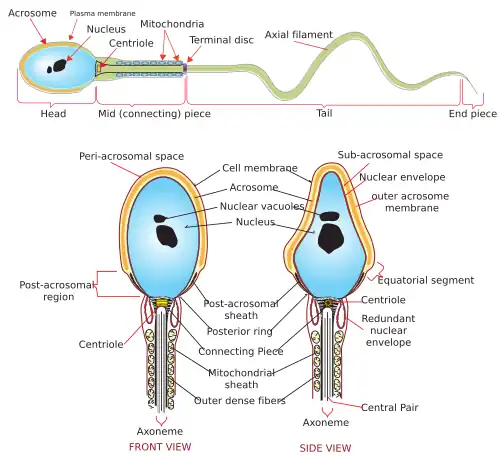
Schematische weergave van de zaadcel met dwarsdoorsneden van de verschillende segmenten. Ax: axoneem, ODF: Buitenste dichte vezel, M: Mitochondriën, LC: Longitudinale kolommen, FS: Vezelige schede, MD: Microtubuli-doublet, CP: Centraal paar microtubuli.
Schema van een menselijke zaadcel met het acrosoom
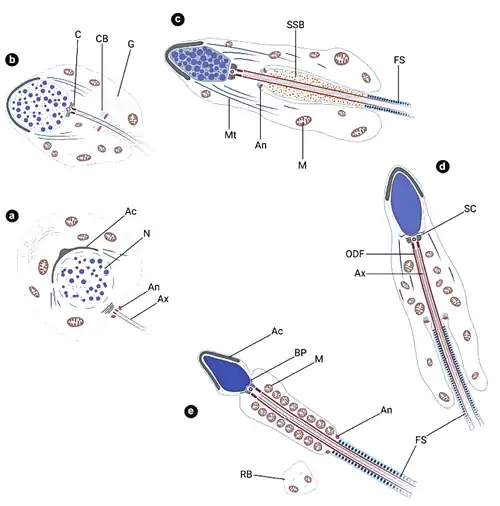
Afbeeldingen vorming van spermatide tot zaadcel (stapsgewijs) bij Galeolaria gemineoa
Stap 1: Het golgicomplex (GC) bevindt zich dicht bij de celkern. Een paar centriolen (C) bevindt zich naast zowel het golgicomplex als het kernmembraan. Bolvormige mitochondriën (M) bevinden zich in clusters in het cytoplasma. (B) Talrijke kleine membraangebonden blaasjes (V) en multivesiculaire lichaampjes (MB) bevinden zich aan het concave oppervlak van het golgicomplex (GC). Centriolen (C) bevinden zich eveneens naast het golgicomplex. Het ruw endoplasmatisch reticulum (RER) is willekeurig verdeeld in het cytoplasma. (C) Bundels microtubuli (Mt) omringen de celkern (N) en vormen een cirkelvormig manchet. M: mitochondriën.
(A) Stap 2 spermatiden zijn ovaal met een bolvormige tot ovale kern (N) met een excentrische nucleolus (Nu) en kleine klontjes chromatine. In de periferie van de kern wordt een duidelijk golgicomplex (GC) waargenomen grenzend aan een bolvormige proacrosomale vacuole (PV) met een elektronendoorlatende matrix, die ontstaat door de fusie van kleine membraangebonden blaasjes. Continue profielen van ruw endoplasmatisch reticulum (RER) lopen rond de kern. Bolvormige mitochondriën (M) zijn willekeurig verspreid door het cytoplasma. (B) De twee centriolen blijven in de buurt van het golgicomplex (GC), maar raken relatief ver van de celkern verwijderd. Terwijl het distale centriool (DC) het celmamembraan nadert, oriënteert het proximale centriool (PC) parallel aan eerstgenoemde en is relatief ver van het celmembraan verwijderd.
Stap 3: (A) Vergelijkbaar met hun directe voorlopers blijven de spermatiden ovaal met een bolvormige tot ovale celkern (N) die staafvormige chromatineblokken bevat. Dikke plekken chromatine hechten zich ook aan het binnenste kernmembraan. Naast het golgicomplex (GC) wordt een bolvormige proacrosomale vacuole (PV) waargenomen die een plek elektronendicht materiaal bevat. Een deel van de mitochondriën (M) transformeert in staafvormige organellen in de spermatiden. (B) Zowel de proximale (PC) als de distale centriolen (DC) blijven in de nabijheid van het golgicomplex (GC) en beginnen zich loodrecht op elkaar te oriënteren. Het ruw endoplasmatisch reticulum (RER) bevindt zich rond de celkern (N). M, mitochondriën. (C) De spermatiden zijn in paren en met elkaar verbonden door een cytoplasmatische brug (CB). Mitochondriën (M) zijn geclusterd in het cytoplasma nabij de cytoplasmatische brug. N, celkern.
Stap 4: (A) Chromatineblokken in de celkern (N) blijven staafvormig, maar worden schaarser dan in voorgaande stappen, wat aangeeft dat chromosoomdecondensatie in dit stadium begint. Mitochondriën (M) en ruw endoplasmatisch reticulum (RER) zijn willekeurig verdeeld over het cytoplasma. (B) De proacrosomale vacuole transformeert tot een proacrosoom (P), waarbij de matrix grotendeels wordt bezet door elektronendicht materiaal. N, celkern. (C) Het distale centriool (DC) naast het golgicomplex (GC) vormt een zweepstaartje (F) dat uit de spermatide steekt. N, celkern; RER, ruw endoplasmatisch reticulum.
Stap 5: (A) Naarmate de nucleaire grootte (N) van de spermatiden afneemt, aggregeren de gedecondenseerde chromosomen tot een retiform patroon. Grote bolvormige mitochondriën (M) worden waargenomen die zich aan één uiteinde van de spermatiden verzamelen. RER: ruw endoplasmatisch reticulum. (B) In het proacrosoom (P) van de spermatiden vormt zich een ringvormige structuur (R) rond de bol (S) in het centrum. M: mitochondriën; N: celkern. (C) De spermatiden blijven met elkaar verbonden door een cytoplasmatische brug (CB). Rondom de celkern bevindt zich ruw endoplasmatisch reticulum (RER).
Stap 6: (A) Een implantatiegroeve (*), een ondiepe verdieping, vormt zich aan het achterste uiteinde van de celkern (N). Het proximale centriool (PC) zit dicht bij de implantatiegroeve, terwijl het distale centriool vast blijft zitten aan het celmembraan. Grote bolvormige mitochondriën (M) migreren naar de pool waar de centriolen zitten. (B) Dwarsdoorsnede stukje proacrosoom, met fragmenten van zowel de ringvormige structuur (R) als de centrale bol (S). De proacrosomale matrix wordt bezet door grove korrels. Een deel van het proacrosoom hecht zich aan het celmembraan. Grote bolvormige mitochondriën zitten naast het proacrosoom. N, celkern. (C) Terwijl het proximale centriool (PC) zich in de implantatiegroeve nestelt, strekt de zweepstaart (F) zich uit vanaf het distale centriool en wordt langer.
Stap 7: (A) Het chromatine in de celkern vertoont een grove korrelvorm en aggregeert tot een groot onregelmatig gevormd blok. Het chromatineblok hecht zich aan het achterste uiteinde van de celkern (N), waar zich een implantatiegroeve (*, witte asterisk) heeft gevormd. Het proximale centriool (PC) blijft dicht bij de groeve en oriënteert loodrecht op het distale centriool (DC). De grote bolvormige mitochondriën (M) worden omgeven door de centriolen en beginnen de mitochondriale mantel aan de basis van de zweepstaart te vormen. (B) Het acrosoom (A) transformeert in een omgekeerde komvorm met het voorste deel stevig aan het celmembraan gehecht. In de acrosomale matrix hechten zowel de centrale bol (S) als de ringvormige structuur (R) zich aan het binnenste achterste membraan van het acrosoom. Het acrosoom ligt naast grote bolvormige mitochondriën (M), die uiteindelijk de mitochondriale omhulling van de zweepstaart vormen. (C) Waar de mitochondriën (M) het celkernmembraan (N) naderen, buigt het celkernmembraan naar binnen en vormt holtes (*, zwarte sterretjes).
Stap 8: (A) Spermatiden zijn nog met elkaar verbonden door een cytoplasmatische brug (CB). De celkern (N) blijft bolvormig, terwijl het chromatineblok transformeert naar een ovale vorm met een zeer gecondenseerd homogeen uiterlijk. Het proximale centriool (PC) strekt zijn microtubuli uit in de implantatiegroeve (*) en oriënteert loodrecht op de zweepstaart (F) die ontspringt uit het distale centriool (DC). Het zich ontwikkelende acrosoom (A) blijft grenzend aan de zweepstaart. (B) De centrale bol hecht zich stevig aan het achterste membraan van het acrosoom (A) en transformeert in een halve bol (Hs). De ringachtige structuur (R), met donkere deeltjes die de binnengrens bekleden, komt ook in direct contact met het acrosomale membraan aan beide zijden. De acrosomale matrix is gevuld met dicht opeengepakte grove korrels. M, mitochondriën; N, celkern. (C) Naarmate de morfogenese van het acrosoom vordert, neemt het volume van de centrale halve bol (Hs) toe en stijgt deze op naar het voorste membraan, wat resulteert in de vorming van een inkeping aan het achterste uiteinde van het acrosoom. N, celkern; R, ringvormige structuur.
Stap 9: A) De spermatiden zijn nog steeds in paren en met elkaar verbonden door een cytoplasmatische brug (CB). De centrale halve bol (Hs) in het acrosoom beslaat de gehele acrosomale matrix en is aan beide zijden verbonden met de ringvormige structuur (R). Het gehele acrosoom vertoont een identieke elektronendichtheid, met uitzondering van de donkere deeltjes die de binnenrand van de ringstructuur bekleden. Meerdere onregelmatig gevormde restlichaampjes (RB) met overtollig cytoplasma verschijnen. M, mitochondriën; N, celkern. (B) Het acrosoom (A) begint te migreren naar het voorste uiteinde van het cytoplasma, dat tegenover de pool ligt waar de zweepstaart (F) en mitochondriën (M) zich bevinden. De mitochondriën blijven stevig vastzitten aan de holtes aan het achterste uiteinde van de celkern (N).
Stap 10: (A) De ovale kern (N) wordt ingenomen door volledig gecondenseerd homogeen granulair chromatine. Het acrosoom (A) bereikt de voorste apex van het cytoplasma, tegenover de pool waar de mitochondriën (M) en zweepostaart zich bevinden. Naarmate de donkere deeltjes zich door de ringachtige structuur verspreiden, lijkt de ring donkerder dan de centrale halve bol (Hs). (B) Vroege stap 10 spermatiden blijven in paren en zijn onderling verbonden door een cytoplasmatische brug (CB). Het cytoplasma bevat clusters van onregelmatig gevormde blaasjes, wat wijst op de continue vorming en uitstoting van restlichaampjes. De vorming van de mitochondriale omhulling wordt voltooid, met vier bolvormige mitochondriën (M) rondom het proximale centriool (PC). *, implantatiegroeve. (C) In late stap 10 spermatiden worden talrijke clusters van onregelmatig gevormde blaasjes gevonden in en grenzend aan het cytoplasma, wat suggereert dat de afvoer van restlichaampjes (RB) in dit stadium dramatisch plaatsvindt. Door de verwijdering van overtollig cytoplasma valt de cytoplasmatische brug die de spermatiden verbindt uiteen, wat resulteert in de vorming van twee onafhankelijke zaadcellen: A, acrosoom; M, mitochondriën; N, celkern.
Zaadcel: (A) Zaadcellen bestaan uit een kapvormig acrosoom (A), een ovale celkern (N), een kort middenstuk met vier bolvormige mitochondriën (M) en een langwerpige zweepstaart (F). Tussen het acrosoom en de celkern vormt zich een subacrosomale ruimte (*, zwarte asterisk), die gelijkmatig verdeeld elektronendicht fibrillair subacrosomaal materiaal bevat. Het proximale centriool (PC) is ingevoegd in de implantatiegroeve (*, witte asterisk) die aan het achterste uiteinde van de celkern is gevormd. De vier mitochondriën nestelen zich in concave holtes aan de achterste celkernrand en omringen de basis van de zweepstaart, waar de annulus (An) wordt gevormd. (B) Een dwarsdoorsnede door de ringstructuur aan de basis van het acrosoom toont de fibrillaire inhoud van de subacrosomale ruimte (*, zwarte asterisk). (C) Een dwarsdoorsnede van het middenstuk toont vier bolvormige mitochondriën (M) die de basis van de zweepstaart omringen. Distaal centriool (DC).
_Cropped_NCR.jpg)