Neuro-epitheliale cel
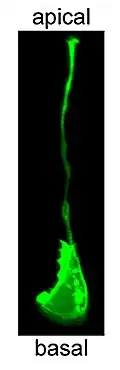

Neuro-epitheliale cellen of neuro-ectodermale cellen zijn afkomstig van het ectoderm en vormen de wand van de gesloten neurale buis in de vroege embryonale ontwikkeling. De neuro-epitheliale cellen zijn met het oppervlak van het zachte hersenvlies en met dat van de vaatholte verbonden door zonula adhaerenscomplexen, waar ze een pseudogestratificeerd epitheel vormen, neuro-epitheel genaamd.[1] Pseudogestratificeerd epitheel is een speciaal type cilindrisch epitheel waarin cellen dun en langwerpig zijn. De celkernen zitten dicht tegen elkaar en dwingt cellen om hun kernen langs de apicobasale as te verdelen, waardoor er meerdere lagen kernen ontstaan binnen een monolaag van cellen, vandaar de term pseudostratificatie.[2] In de embryonale proliferatiefase vormt het neuro-epitheel (ook wel embryonaal neuro-epitheel genoemd, om het te onderscheiden van dat aanwezig in volwassen organismen) of neuro-ectoderm de matrix met neurale stamcellen.
Interfase celkernbewegingen
Een kenmerkende eigenschap van pseudogestratificeerd epitheel is de gecoördineerde bewegingen van celkernen tijdens de celcyclus, Interfase celkernbewegingen genoemd. Interfase celkernbewegingen bestaan uit verschillende stappen: een apicale naar basale beweging die plaatsvindt tijdens de G1- en S-fasen van de celcyclus en een basale naar apicale celkern beweging die plaatsvindt tijdens de G2- en M-fasen. De apicale-waartse beweging, soms aangeduid als PRAM (Pre-mitotic Rapid Apical Movement), kan worden bereikt via microtubuli zoals in het neuro-epitheel van kuikens of in de hersenen van muizen- en rattenembryo's en ook in het netvlies van pasgeboren muizen. Dergelijke bewegingen kunnen ook op een actomyosine-afhankelijke manier plaatsvinden, zoals waargenomen in het netvlies van vissenembryo's. De terugkeer van celkernen naar basale posities na mitotische celdelingen werd aanvankelijk voorgesteld als passief en een direct gevolg van celkern gedrang in de apicale regio van pseudogestratificeerd epitheel. Er zijn echter talrijke bewijzen die wijzen op een actieve rol van het cytoskelet bij celkernverplaatsing van apicaal naar basaal. Kif1A, een anterograde moleculaire motor van microtubuli, is bijvoorbeeld vereist voor de beweging van celkernen in rattenhersenen van apicaal naar basaal. Anterograde transport is de beweging van moleculen/organellen in het cellichaam naar het celmembraan. Bovendien is bij de grote hersenen van muizen aangetoond dat myosine II essentieel is voor de beweging van apicaal naar basaal. Verder zijn in frettenhersenen de bewegingen van celkernen van apicaal naar basaal sneller dan de bewegingen van basaal naar apicaal, wat suggereert dat de celkernbeweging naar basale hersengebieden actief is. De tegenovergestelde beweging wordt echter waargenomen bij muizen. Al deze observaties geven aan dat Interfase celkernbewegingen worden gereguleerd door cytoskelet-afhankelijke mechanismen en dat het daadwerkelijk gebruikte mechanisme verschilt van soort tot soort en van orgaan tot orgaan.[2]
Embryonale neurale ontwikkeling
Ontwikkeling van de hersen
Tijdens de derde week van de embryonale ontwikkeling ( gerekend vanaf de bevruchting) beginnen de hersenen in het embryo zich te ontwikkelen in een proces dat morfogenese wordt genoemd.[3] Neuro-epitheliale cellen van het ectoderm beginnen zich snel te vermenigvuldigen en vouwen zich om de neurale plaat te vormen, die in de vierde week van de embryonale groei invagineert en de neurale buis vormt.[3] De vorming van de neurale buis polariseert de neuro-epitheliale cellen door de apicale zijde van de cel naar binnen te richten, wat later de ventriculaire zone wordt, en de basale zijde is naar buiten gericht, wat contact maakt met het zachte hersenvlies, het buitenste oppervlak van de zich ontwikkelende hersenen.[4] Als onderdeel van deze polariteit brengen neuro-epitheliale cellen prominine-1 tot expressie in het apicale celmembraan en in zonula occludens om de celpolariteit te behouden.[5] Integrine alfa 6 verankert de neuro-epitheliale cellen aan het basaal membraan.[5] De neurale buis begint als een enkele laag epitheelcellen, maar snelle proliferatie van neuro-epitheliale cellen creëert extra lagen, het meerrijig epitheel, en uiteindelijk drie verschillende groeigebieden.[3][5] Omdat deze extra lagen de apicale-basale polariteit vormen, moet deze worden downgereguleerd.[4] Verdere proliferatie van de cellen in deze gebieden leidt tot drie verschillende gebieden in de hersenen: de voorhersenen, de middenhersenen en de achterhersenen. De neurale buis geeft ook aanleiding tot het ruggenmerg.[3]
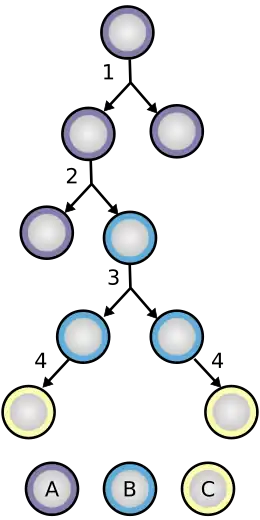
Proliferatie van neuro-epitheliale cellen
Neuro-epitheliale cellen zijn een type stamcellen en hebben het vermogen om zichzelf te vernieuwen. Tijdens de vorming van de neurale buis ondergaan neuro-epitheliale cellen symmetrische delingen die aanleiding geven tot twee nieuwe neuro-epitheliale cellen. In een later stadium van de ontwikkeling van de hersenen beginnen neuro-epitheliale cellen zichzelf te vernieuwen en vormen door asymmetrische deling niet-stamcelvoorlopers, zoals radiale gliacellen. Expressie van Tis21, een antiproliferatief gen, zorgt ervoor dat de neuro-epitheliale cel de overstap maakt van proliferatieve deling naar neuronische deling. Veel van de neuro-epitheliale cellen delen zich ook in radiale gliacellen, een vergelijkbare, maar meer lotbeperkte cel. Omdat het een cel is met een beperkter lot, zal de radiale gliacel postmitotische zenuwcellen, intermediaire voorlopercellen of astrocyten genereren in de gliogenese. Tijdens de neuro-epitheliale celdeling zorgt interkinetische celkern migratie ervoor dat de cellen zich onbeperkt kunnen delen terwijl ze een dichte pakking behouden. Tijdens G1 migreert de celkern naar de basale zijde van de cel en blijft daar voor de S-fase en migreert naar de apicale zijde voor de G2-fase. Deze migratie vereist de hulp van microtubuli en actinefilamenten.[5]
Radiale gliacelvorming
Neuro-epitheliale cellen vormen radiale gliale voorlopercellen in de vroege embryonale ontwikkeling. Om deze verandering te bewerkstelligen, beginnen neuro-epitheliale cellen hun epitheliale kenmerken te downreguleren door de expressie van occludine, een zonula occludens-eiwit, te stoppen.[4] Verlies van occludine veroorzaakt een verlies van de eerdere zonula occludens-afdichtingen die nodig zijn voor de generatie van neuroblasten. Een ander zonula occludens-eiwit, PARD3, blijft aan de apicale zijde van de cel co-lokaliserend met N-cadherine en houdt het apicale vlak van de neuro-epitheliale cel intact.[5] Bij afwezigheid van occludine gaat er nog steeds wat polariteit verloren en vormt de neuro-epitheliale cel de radiale gliacel.[5]
 |
- voetnoten
- ↑ Sadler, T (2006). Langman's medical embryology., 11th.. Lippincott William & Wilkins, 295–299. ISBN 9780781790697.
- ↑ a b Ferreira MA, Despin-Guitard E, Duarte F, Degond P, Theveneau E (2019) Interkinetic nuclear movements promote apical expansion in pseudostratified epithelia at the expense of apicobasal elongation. PLoS Comput Biol 15(12): e1007171. https://doi.org/10.1371/journal.pcbi.1007171
- ↑ a b c d McDonald, A. (2007). Prenatal Development - The Dana Guide. The Dana Foundation. ISBN 978-1-932594-10-2. Gearchiveerd op 8 januari 2022. Geraadpleegd op 7 december 2011.
- ↑ a b c Zolessi, F. R. (2009). Encyclopedia of Life Sciences. DOI:10.1002/9780470015902.a0000826.pub2, "Vertebrate Neurogenesis: Cell Polarity". ISBN 978-0470016176.
- ↑ a b c d e f Götz, M., Huttner, W. B. (2005). The cell biology of neurogenesis. Nature Reviews Molecular Cell Biology 6 (10): 777–788. PMID 16314867. DOI: 10.1038/nrm1739.
- bronvermelding
- Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Neuroepithelial cell op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar.