Unipolaire borstelcel
Unipolaire borstelcellen zijn een klasse van exciterende glutamatergische interneuronen die zich bevinden in de granulaire laag van de kleine hersenencortex en ook in het granulaire celdomein van de cochleakern.[1][2][3] De cochleakernen bevinden zich aan de dorso-laterale zijde van de hersenstam en overspannen de grens tussen de pons en de medulla oblongata.
Er worden twee typen unipolaire borstelcellen onderscheiden. Type I vormt calretinine (een calciumbindend eiwit) en type 2 vormt mGluR1α (een G-proteïnegekoppelde receptor uit de C klasse).[4]
Structuur
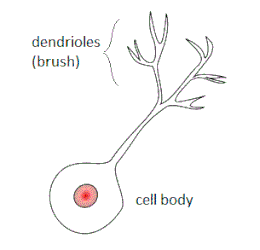
De unipolaire borstelcel heeft een rond of ovaal cellichaam met meestal één korte dendriet die eindigt in borstelachtige, korte dendriolen (dendrieten die uniek zijn voor unipolaire borstelcellen). Deze borsteldendriolen vormen zeer grote synaptische verbindingen. De dendritische borstel en de grote uiteinden van de axonuitlopers zijn betrokken bij de vorming van cerebellaire glomeruli. De cerebellaire glomerulus is een kleine, verstrengelde massa zenuwvezeluiteinden in de granulaire laag van de klein hersenencortex. Deze bestaat uit postsynaptische korrelceldendrieten en presynaptische uiteinden van mosvezels.[5]
De unipolaire borstelcel heeft één korte dendriet waar de korrelcel er vier of vijf heeft.
De borsteldendriolen vormen talrijke, dunne uitlopers, filopodia genaamd, die uniek zijn voor unipolaire borstelcellen. De filopodia komen uit de hele zenuwcel, zelfs vanuit de dendritische stam en het cellichaam in sommige cellen. Hoewel unipolaire borstelcelfilopodia geen synaptische verbindingen hebben, zijn ze desalniettemin betrokken bij celsignalering.[6]
Functie
Unipolaire borstelcellen zijn intrinsiek vurende zenuwcellen en worden beschouwd als een klasse van exciterende "lokale circuitneuronen".[7] Ze werken samen met vestibulaire vezels om signalen te integreren die te maken hebben met de oriëntatie van het hoofd die reflexgedrag moduleert.[8] Unipolaire borstelcellen versterken input van de vestibulaire ganglia en kernen door de excitatie binnen de korrelige laag te verspreiden en te verlengen.[2][9] Ze ontvangen glutamatergische input op hun dendritische borstel van een enkel mosvezeluiteinde in de vorm van een gigantische glutamatergische synaps en maken glutamatergische synapsen met korrelcellen en andere unipolaire borstelcellen.
Locatie
Unipolaire borstelcellen zijn talrijk in die gebieden die verband houden met vestibulaire functies. Bij zoogdieren vertonen unipolaire borstelcellen een ongelijkmatige verdeling binnen de korrelceldomeinen van het metencephalon, met de grootste dichtheid in de vermis, onderdeel van het flocculus/paraflocculus-complex en de lagen 2-4 van de dorsale cochleakern.[7] In de kleine hersenen van ratten overtreffen unipolaire borstelcellen het aantal golgicellen met een factor 3 en zijn ze ongeveer gelijk aan het aantal purkinjecellen. Net als andere glutamatergische cellen van de kleine hersenen ontstaan unipolaire borstelcellen in de rhombische lip.[10] De rhombische lip is een achterste gedeelte van het zich ontwikkelende metencephalon dat tijdelijk kan worden herkend in het embryo van gewervelden.
Geschiedenis
Unipolaire borstelcellen werden voor het eerst beschreven in 1977 door Altman en Bayer, die ze "bleke cellen" noemden.[11] De term "unipolaire borstelcel" werd voor het eerst geïntroduceerd begin jaren negentig, waarbij bleekcellen, Rat-302-cellen, monodendritische cellen, kastanjecellen en mittcellen onder dezelfde naam werden heringedeeld. Het Federative International Committee on Anatomical Terminology (FICAT), een subcommissie van de International Federation of Associations of Anatomists (IFAA), erkende de "unipolaire borstelcel" officieel als een nieuw celtype van de kleine hersenencortex in 2008.[7]
Klinische betekenis
Unipolaire borstelcellen in lobule VII van de kleine hersenen worden in sommige gevallen door de ziekte van Pick aangetast, waarbij ze cytoskeletafwijkingen ontwikkelen en herkend worden door antilichamen tegen abnormaal gehyperfosforyleerde tau-eiwitten.[12] Unipolaire borstelcellen zijn ook in verband gebracht met de disfunctie van evenwicht en motorische coördinatie die aanwezig is bij het syndroom van Down.[13]
- Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Unipolar brush cell op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar.
- ↑ Mugnaini, Enrico, Sekerková, Gabriella, Martina, Marco (5 november 2010). The unipolar brush cell: a remarkable neuron finally receiving deserved attention 66 (1–2): 220–245. PMID 20937306. PMC 3030675. DOI: 10.1016/J.BRAINRESREV.2010.10.001.
- ↑ a b Englund, C. (2006). Unipolar Brush Cells of the Cerebellum Are Produced in the Rhombic Lip and Migrate through Developing White Matter. Journal of Neuroscience 26 (36): 9184–9195. ISSN: 0270-6474. PMID 16957075. PMC 6674506. DOI: 10.1523/JNEUROSCI.1610-06.2006.
- ↑ Diño, M.R, Schuerger, R.J, Liu, Y.-B, Slater, N.T, Mugnaini, E (2000). Unipolar brush cell: a potential feedforward excitatory interneuron of the cerebellum. Neuroscience 98 (4): 625–636. ISSN: 0306-4522. PMID 10891606. DOI: 10.1016/S0306-4522(00)00123-8.
- ↑ McDonough A, Elsen GE, Daza RM, Bachleda AR, Pizzo D, DelleTorri OM and Hevner RF (2021) Unipolar (Dendritic) Brush Cells Are Morphologically Complex and Require Tbr2 for Differentiation and Migration. Front. Neurosci. 14:598548. doi: 10.3389/fnins.2020.598548
- ↑ Cerebellar glomerulus. NeuroLex. The Neuroscience Information Framework (14 Oct 2011). Geraadpleegd op 24 June 2014.
- ↑ Jaarsma, D, Diño, MR, Ohishi, H, Shigemoto, R, Mugnaini, E (1998). Metabotropic glutamate receptors are associated with non-synaptic appendages of unipolar brush cells in rat cerebellar cortex and cochlear nuclear complex. J Neurocytol 27 (5): 303–27. PMID 9923978. DOI: 10.1023/A:1006982023657.
- ↑ a b c Mugnaini, E, Sekerková, G, Martina, M (2011). The unipolar brush cell: a remarkable neuron finally receiving deserved attention. Brain Res Rev 66 (1–2): 220–45. PMID 20937306. PMC 3030675. DOI: 10.1016/j.brainresrev.2010.10.001.
- ↑ Harris J, Moreno S, Shaw G, Mugnaini E (December 1993). Unusual neurofilament composition in cerebellar unipolar brush neurons. Journal of Neurocytology 22 (12): 1039–59. PMID 8106879. DOI: 10.1007/BF01235748.
- ↑ Diño, Maria, Perachio, Adrian, Mugnaini, Enrico (2001). Cerebellar unipolar brush cells are targets of primary vestibular afferents: an experimental study in the gerbil. Experimental Brain Research 140 (2): 162–170. ISSN: 0014-4819. PMID 11521148. DOI: 10.1007/s002210100790.
- ↑ Englund C, Kowalczyk T, Daza RA, Dagan A, Lau C, Rose MF, Hevner RF (2006). Unipolar brush cells of the cerebellum are produced in the rhombic lip and migrate through developing white matter.. J. Neurosci. 26 (36): 9184–95. PMID 16957075. PMC 6674506. DOI: 10.1523/JNEUROSCI.1610-06.2006.
- ↑ Altman J, Bayer SA (1977). Time of origin and distribution of a new cell type in the rat cerebellar cortex.. Exp Brain Res 29 (2): 265–74. PMID 913518. DOI: 10.1007/bf00237046.
- ↑ Braak E, Arai K, Braak H (1999). Cerebellar involvement in Pick's disease: affliction of mossy fibers, monodendritic brush cells, and dentate projection neurons.. Exp Neurol 159 (1): 153–63. PMID 10486184. DOI: 10.1006/exnr.1999.7131.
- ↑ Harashima C, Jacobowitz DM, Stoffel M, et al. Elevated expression of the G-protein-activated inwardly rectifying potassium channel 2 (GIRK2) in cerebellar unipolar brush cells of a Down syndrome mouse model. Cell Mol Neurobiol. 2006;26(4-6):719-34.